
和歌山県田辺湾口「沖島」の大サンゴ群集 filmed2010 <吉野熊野国立公園田辺白浜海域公園地区>
生物多様性モニタリング調査風景<田辺白浜海域公園地区>
















生物多様性モニタリングReport
2024年Report
2024年度事業は、公益財団法人公益推進協会「自然公園等保護基金」、並びに独自調査により実施しました。
🔲事業内容
1.生物多様性の保全と調査(田辺白浜海域公園地区)
サンゴ被度・優占種・稚サンゴ・白化、魚類や無脊椎動物、底質構成、サンゴ捕食動物と被害状況等を調査。海底にラインを引き、そのラインに沿って対象種等を目視で数え数値化。
2.オオカワリギンチャクの生息状況調査(田辺湾ショウガセ海域公園地区)
1mの方形枠を設置し写真を撮影、その写真を解析し個体数を集計。水温の推移を捉えるため、連続水温計ロガーを設置し長期のモニタリングを実施。➡オオカワリギンチャク・タコアシサンゴ頁を参照下さい。
3.サンゴを食害するレイシ貝類の除去(田辺市天神崎海域)
サンゴが白化する箇所を探し、サンゴの枝中から、ピンセットを用い、レイシ貝類をつまみ採り、サイズ別に集計。➡レイシ貝類調査・除去を参照下さい。
🔲結果
1.田辺・白浜海域の生物多様性調査と水温計ロガー設置
(1)サンゴ群集の再生状況の調査(ミドリイシ属のサンゴのみ対象)
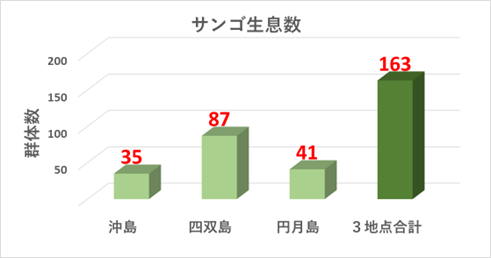
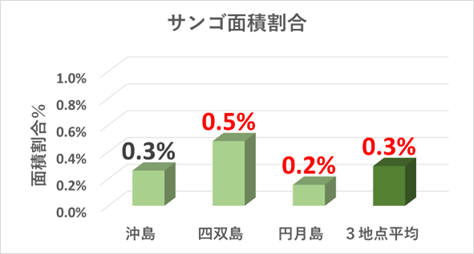
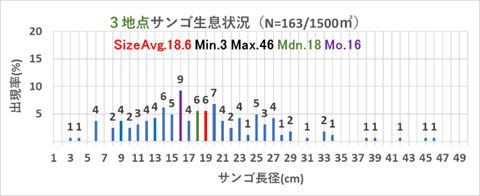
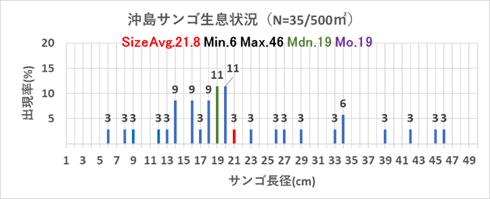
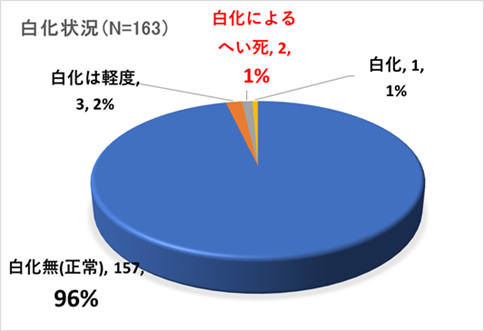
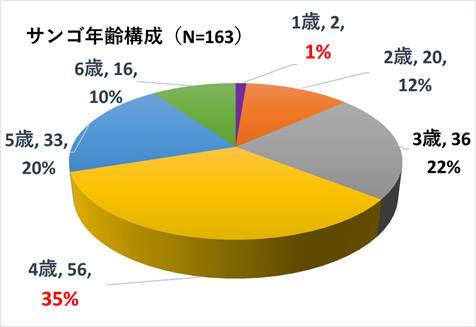
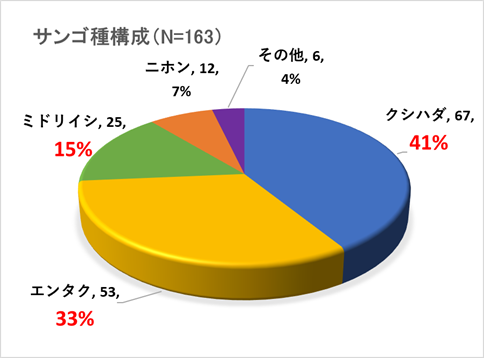
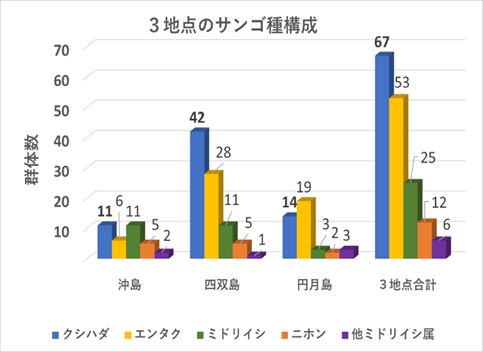
➢田辺白浜海域公園地区のミドリイシ属サンゴ群集の消失
2018年冬季の低水温ショック(32年ぶりの大寒波と黒潮大蛇行の要因により、サンゴの耐低水温限界とされる15℃以下を2か月間継続した。)により、サンゴ群集全域に大規模な白化現象が発生し、壊滅的な被害を受けた。本海域の重要なサンゴ群生地であった沖島・四双島・円月島の高被度サンゴ群集においても、ミドリイシ属サンゴの生残は確認できなかった。
➢新規加入のミドリイシ属サンゴを多数確認
本年度調査により、ミドリイシ属稚サンゴの成長を多数確認できた。このことは、本海域以南の串本や沖縄等のサンゴの産卵により、幼生が流れ着き(新規加入)、成長していることの証明となった。
➢サンゴ群集の再生状況の調査の継続必要
サンゴ新規加入の多寡とその成長を捉えることができた本調査は、大いに有意義であった。しかしながら、総調査面積は1500㎡であり、本海域全体の標本とするのにはあまりにも範囲が狭い。よって、来期以降も本調査を継続し、知見を蓄積しつつ評価することが重点課題となる。
➢水温計測モニタリング継続
本年度から新たに連続水温計(1時間毎計測)を設置したので、今後は水温変動による影響も含めたサンゴの再生過程のモニタリングが可能。次年度に回収(新たな水温計と交換)し年間の水温変動を捉える。
2.田辺・白浜海域の生物多様性調査
(1)底質構成(全種サンゴを対象)
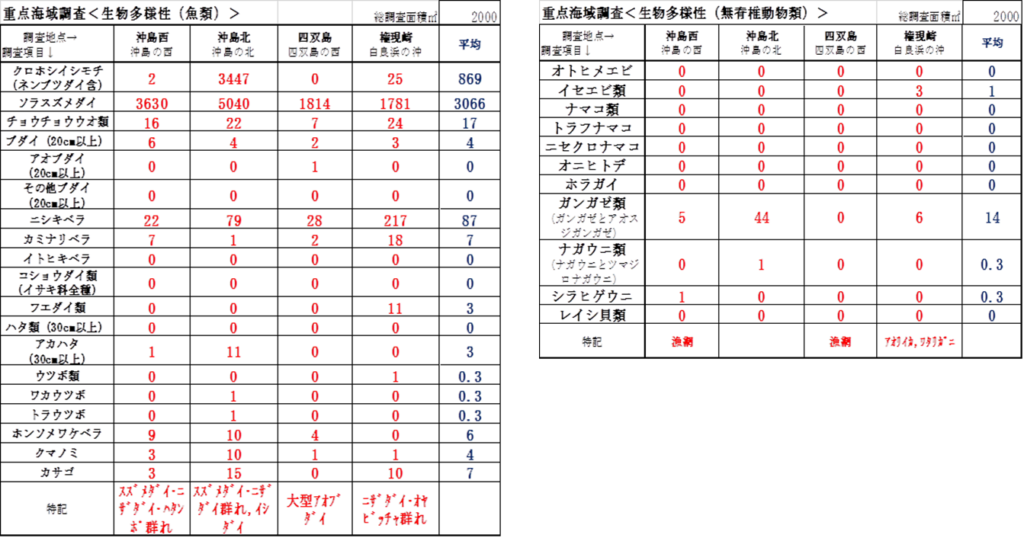
(2)魚類
(3)無脊椎動物
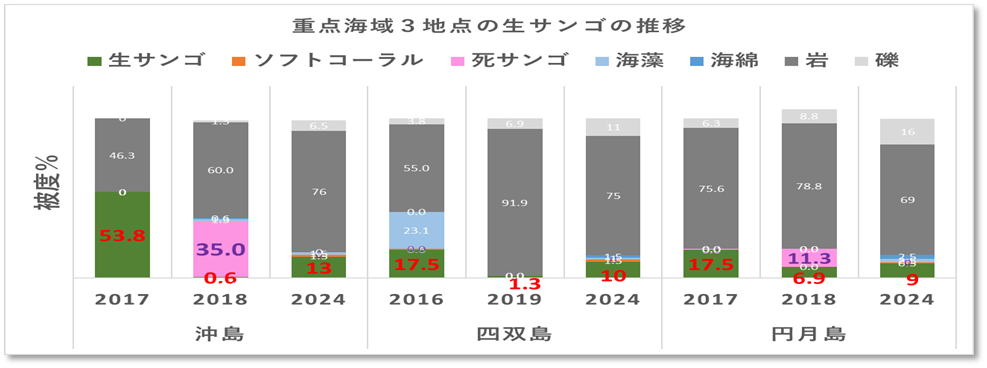
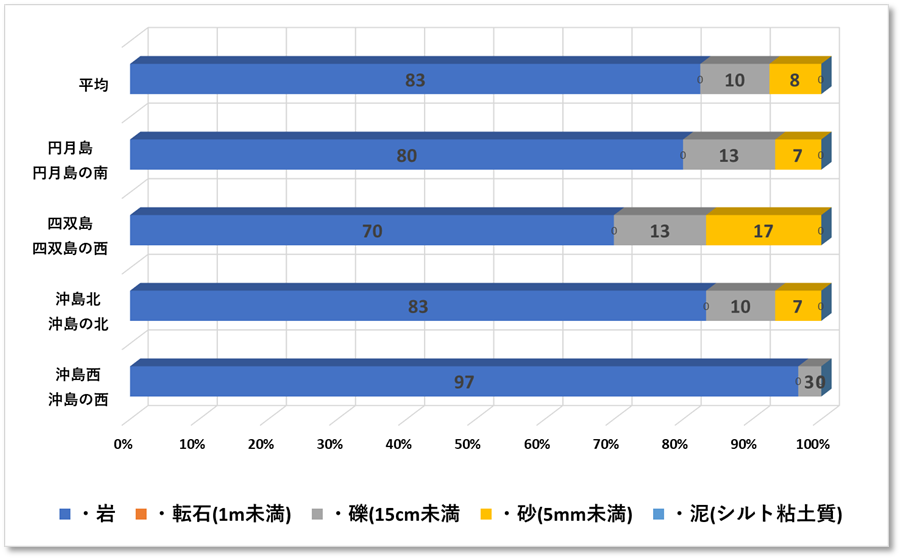
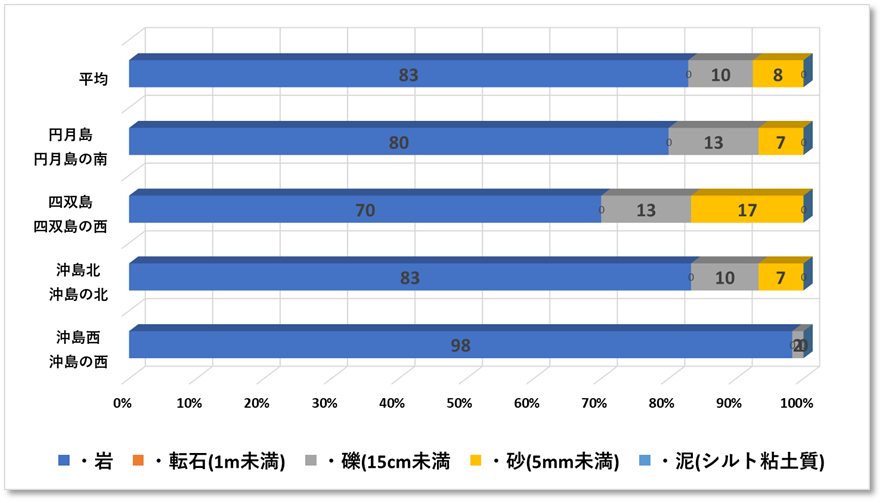
➢底質構成
重点海域の3地点(沖島・四双島・円月島)の生サンゴについてみると、2018年冬季の低水温ショック時に大きく減少したが、本年度調査では微増加している。特に沖島は13%と再生傾向が顕著。
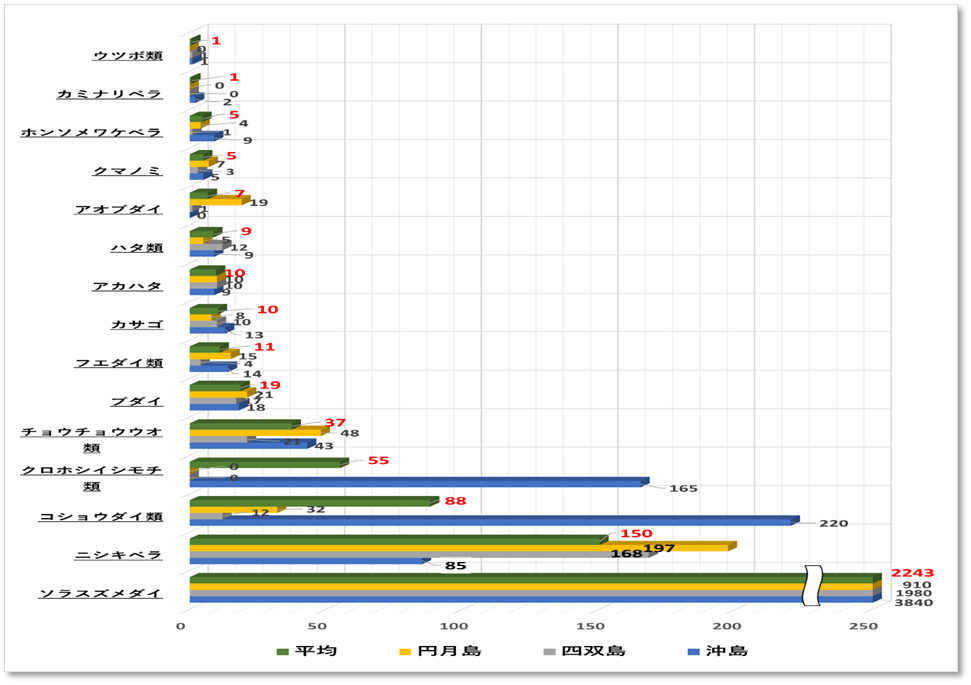
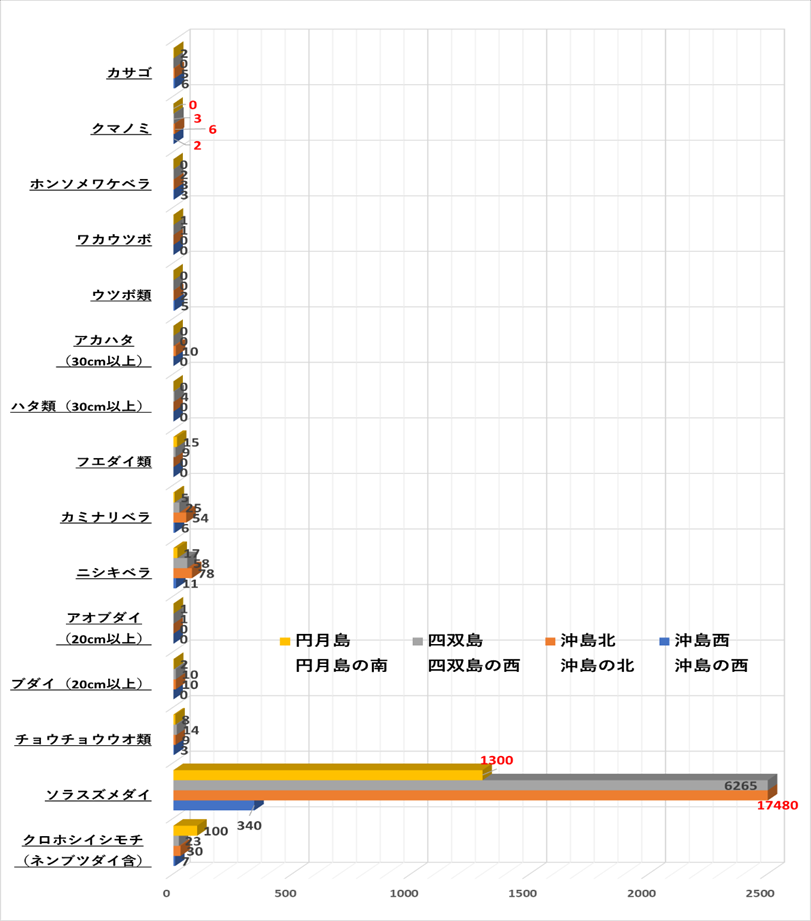
➢魚類
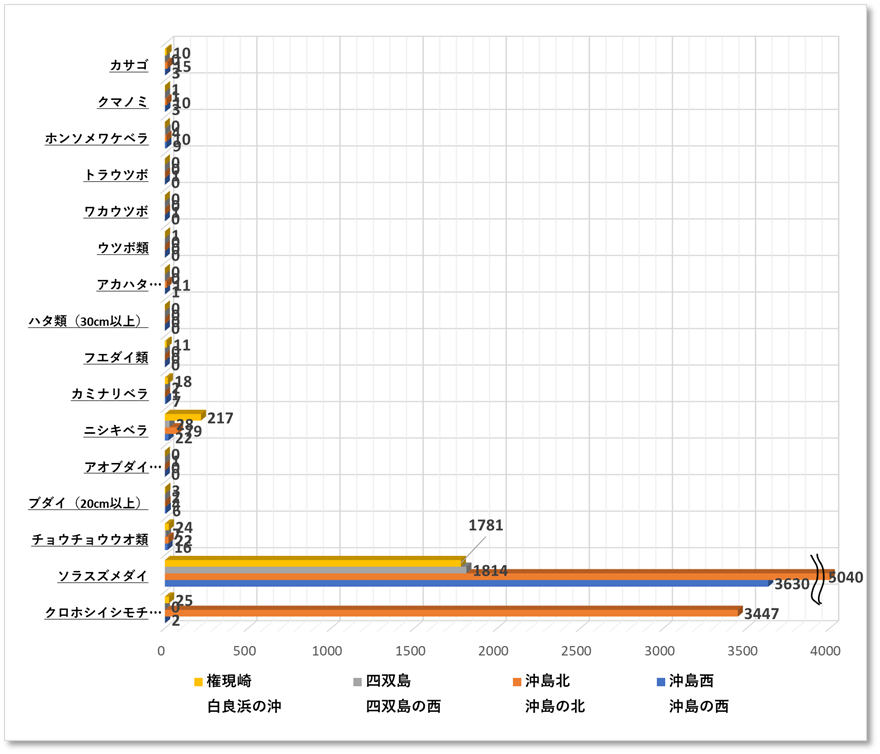
優占種はソラスズメダイが最も多く、次にニシキベラ、コショウダイ類(主はイサキ)イシモチ類と続く。
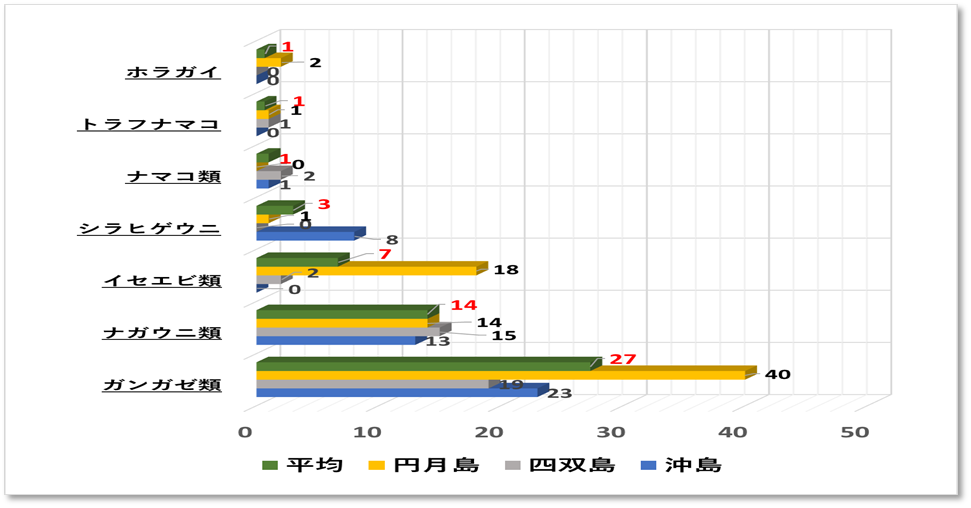
➢無脊椎動物
優占種はカンガゼ類が最多であり、次にナガウニ類、イセエビ類と続く。サンゴを食害するオニヒトデは確認できなった。またレイシ貝類は10個前後の集団が点在する場所もあったが、問題となるような生息数ではなかった。
➢生物多様性調査の継続
今後は、サンゴ群集の再生状況や健康度合い、レイシ貝類やオニヒトデ等のかく乱要因、人為的影響等の調査を継続し、①科学的なデータを蓄積。②モニタリングによる変化を捉え、保全に繋げる。








2021年~2019年結果報告
本調査は里地・里山環境保全推進事業(和歌山県自然環境室)並びに独自調査により実施。
業務対象地域:和歌山県田辺市および白浜町沿岸海域のサンゴ群集浅海域294ha、ならびに深海域1ha
※深海域についてはオオカワリギンチャク・タコアシサンゴ頁を参照下さい。
🔲活動位置図
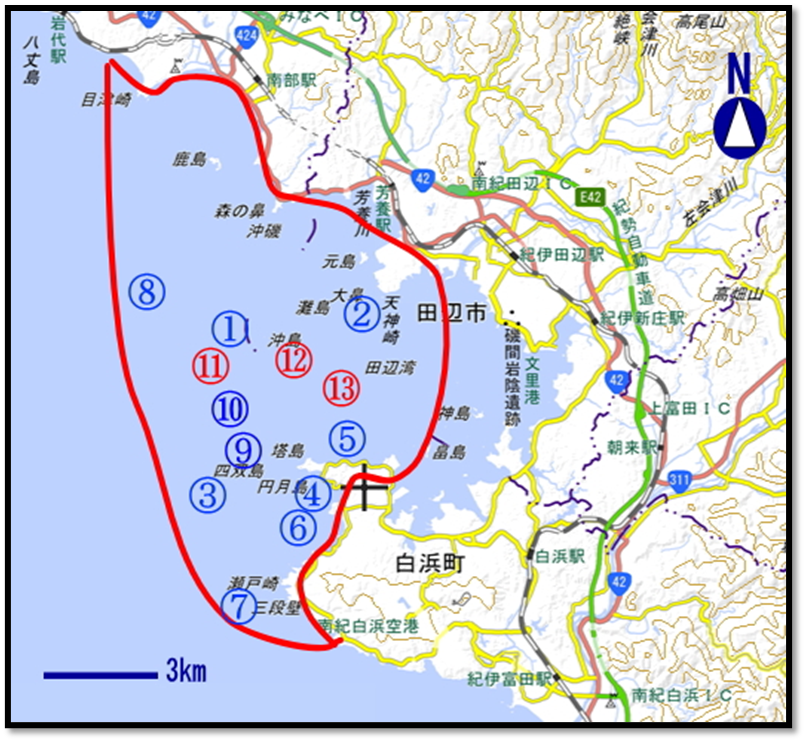
出典:国土地理院の地理院地図を加工して作成
| 海域区分 | 代表する地域名称、 (ダイビングポイント名称等) | 面積 (海図等よりおよその面積を算出) |
| サンゴ群集海域 総面積294ha | ①沖島 | 1000m×1000m = 100 ha |
| ②天神崎 | 50m×500m = 25 ha | |
| ③四双島 | 500m×500m = 25 ha | |
| ④円月島 | 1000m×300m = 30 ha | |
| ⑤江津良 | 1000m×300m = 30 ha | |
| ⑥権現崎 | 1000m×300m = 30 ha | |
| ⑦三段壁 | 1000m×300m = 30 ha | |
| ⑨ エドセ | 100m×100m = 1 ha | |
| ⑩ 中島 | 300m×300m = 9 ha | |
| ⑪ ロングアーチ | 200m×200m = 4ha | |
| ⑫ 沖島東 | 200m×200m = 4 ha | |
| ⑬ 中ノ島 | 200m×300m = 6 ha | |
| 深海域 | ⑧ショウガセ | 100m×100m = 1 ha |
| 集計 | 13地点 | 295 ha |
🔲令和元年(2019年)重点海域調査


〇サンゴの生息状況 4地点の生サンゴ被度は1%程度であり、白化したサンゴや1年以内にへい死したサンゴはみつからなかった。サンゴ優占型形状は塊状のキクメイシ類、それ以外はトゲトサカなどのソフトコーラルであった。
表Ⅲ.8.1 令和元年度 ミドリイシ属の稚サンゴ出現数
| 稚サンゴサイズ(cm長径) | 沖島西 | 沖島北 | 四双島 | 権現崎 |
| 5未満 | 0 | 1 | 7 | 0 |
| 5~10未満 | 2 | 0 | 8 | 0 |
| 合計 | 2 | 1 | 15 | 0 |
〇沖島西 島の西側地先に位置し、陸地(最大でも海抜数メートル)から、水底の平坦地である水深10数メートルまで最初はゆるやかに、水深6mをこえたあたりから傾斜は急こう配になっていた。傾斜域は全て岩礁で構成されていた。また、水底の平坦地は礫・転石・砂で構成されていた。本調査は傾斜域のみで調査したので、岩100%であり多様性に乏しい結果となった。。
〇沖島北 沖島の北側域で、干潮時に洗岩が現れる程度の見印しかない。水中は0mから、水底の平坦地である水深10数メートルまで傾斜する。傾斜域は全て岩礁で構成されていた。また、水底の平坦地は転石・礫・砂で構成されていた。
〇四双島 島の西側地先で、陸地(最大でも海抜数メートル)から、水底の平坦地である水深10数メートルまで傾斜する。傾斜域は岩礁・転石・礫・砂で構成され、西側から島に向かい水路が多く存在した。アーチや横穴、ポットホールが多く、変化に富む地形であった。
〇権現崎 権現崎は白良浜の沖600mに位置し、干潮時に現れる洗岩は南西に伸長し、水中は0mから、水底の平坦地である水深10mまで傾斜する。傾斜域は岩礁・転石・礫・砂で構成され、比較的単調な地形であった。
〇魚類で最も多様性に富む地点
沖島北は魚類の種類と数が多く、他地点よりも多様性に富んでいた。調査対象種以外ではスズメダイ・ニザダイ・ハタンポの群れを確認した。
〇大型定着性魚類 優占種はブダイであった。次に多くみつかったのはアカハタであった。大型種としては四双島で大型のアオブダイを確認した。
〇無脊椎動物 ガンガゼ類がもっとも多くみつかった。権現崎ではタワシウニを多く確認した。調査対象種以外ではアオリイカ十数匹の小集団やワタリガニを確認した。
〇表在底性生物 優占種はトゲトサカやユビノウトサカなどのソフトコーラルであった。権現崎ではタワシウニを多く確認した。
〇植被優占 優占種はヒバやカニノテなどの石灰藻類であった。水産上の重要種であるカジメ・ワカメ・ホンダワラ類などは衰退期ゆえに確認できなかった。
〇特記
四双島では真新しい漁網が散見できた。漁船の転覆事故による漁網の流出と考えられる。沖島西の漁網はエビ網であった。
🔲令和2年(2020年)重点海域調査


〇サンゴの生息状況
4地点の生サンゴ被度は1%程度であり、白化したサンゴや1年以 内へい死したサンゴはみつからなかった。サンゴ優占型形状は塊状のキクメイシ類、それ以外はトゲトサカ等のソフトコーラルであった。ミドリイシ属稚サンゴのサイズと個数を以下に示す。
表Ⅲ.8.2 令和2年度 ミドリイシ属の稚サンゴ出現数
| 稚サンゴサイズ(cm長径) | 沖島西 | 沖島北 | 四双島 | 円月島 |
| 5未満 | 6 | 2 | 8 | 8 |
| 5~10未満 | 9 | 10 | 23 | 25 |
| 10~15未満 | 2 | 2 | 0 | 3 |
| 合計 | 17 | 14 | 31 | 36 |
〇円月島 円月島の南側地先で、島から平坦地の水深10数メートルまで、岩礁・転石・礫・砂・アーチ・穴・ポットホール・水路など多く確認でき、変化に富む地形であった。
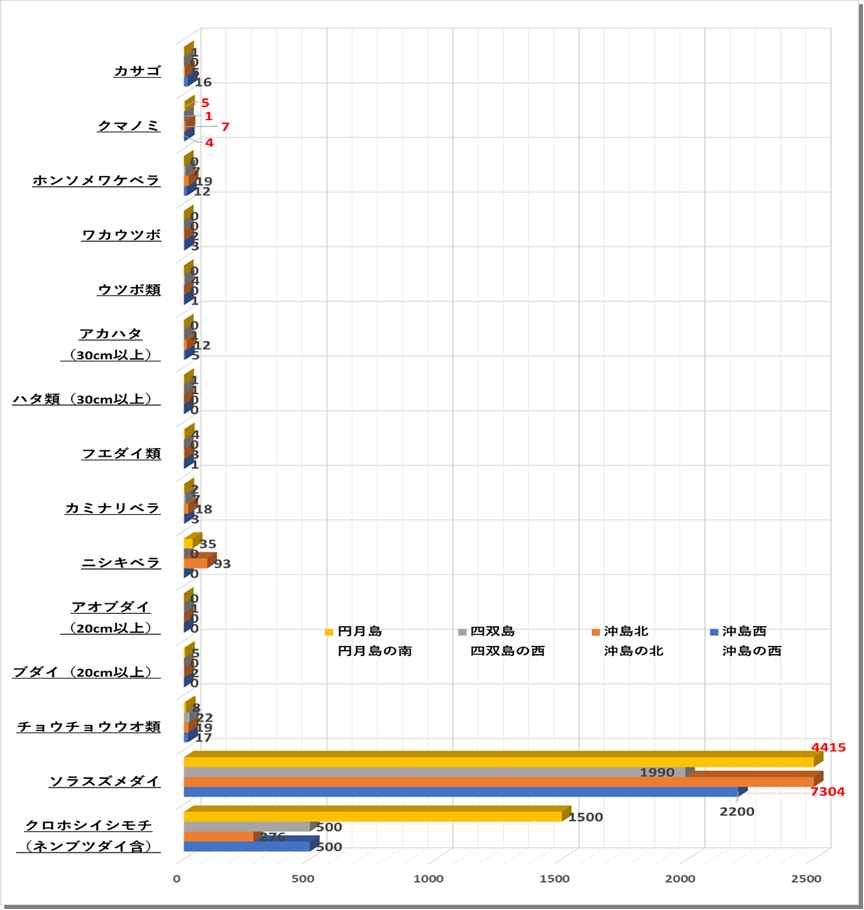
〇魚類 4地点の平均値では、ソラスズメダイ3977匹、次いでクロホシイシモチ694匹、ニシキベラ32匹、ホンソメワケベラ10匹であった。クマノミの平均値は4匹、成魚となりペアで卵を育てる様子を確認した。
〇大型定着性魚類 優占種はブダイであった。四双島と円月島では大型のアオブダイを確認した。
〇無脊椎動物類 ガンガゼ類がもっとも多くみつかった。
〇表在底性生物 優占種はトゲトサカやユビノウトサカなどのソフトコーラルであった。
〇植被優占 優占種はヒバやカニノテなどの石灰藻類であった。水産上の重要種であるカジメ・ワカメ・ホンダワラ類などは衰退期ゆえに確認できなかった。アミジグサ類も多く確認した。
〇特記 四双島と沖島西では漁網が岩等に絡みつく様子が目立った。
🔲令和3年(2021年)重点海域調査


〇サンゴの生息状況
4地点の生サンゴ被度は1%程度であり、白化したサンゴや1年以内へい死したサンゴはみつからなかった。サンゴ優占型形状は塊状のキクメイシ類、それ以外はトゲトサカ等のソフトコーラルであった。ミドリイシ属稚サンゴのサイズと個数を以下に示す。
表Ⅲ.8.3 令和3年度 ミドリイシ属の稚サンゴ出現数
| 稚サンゴサイズ(cm長径) | 沖島西 | 沖島北 | 四双島 | 円月島 |
| 5未満 | 18 | 5 | 6 | 3 |
| 5~10未満 | 24 | 14 | 30 | 13 |
| 10~15未満 | 19 | 12 | 33 | 13 |
| 15~20未満 | 2 | 2 | 1 | 1 |
| 20~25未満 | 0 | 0 | 0 | 2 |
| 25~30未満 | 0 | 0 | 0 | 1 |
| 合計 | 63 | 33 | 70 | 33 |
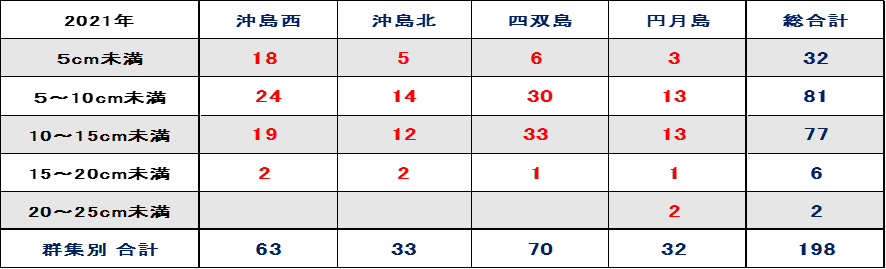
稚サンゴ種はクシハダミドリイシ・エンタクミドリイシ・ニホンミドリイシ・ミドリイシの4種と暫定した。
〇大型定着性魚類 優占種はブダイであった。四双島と円月島では大型のアオブダイを確認した。
〇植被優占 優占種はヒバやカニノテなどの石灰藻類であった。水産上の重要種であるカジメ・ワカメ・ホンダワラ類などは衰退期ゆえに確認できなかった。アミジグサ類も多く確認した。
〇魚類 4地点の平均値では、ソラスズメダイ6346匹、次いでニシキベラ41匹、クロホシイシモチ40匹、カミナリベラ23匹であった。 クマノミの平均値は3匹、成魚となりペアで卵を育てる様子を確認した。
〇無脊椎動物類 ナガウニ類がもっとも多くみつかった。
〇特記 四双島と沖島西では漁網が岩等に絡みつく様子が目立った。
🔲令和元年(2019年)被害状況調査

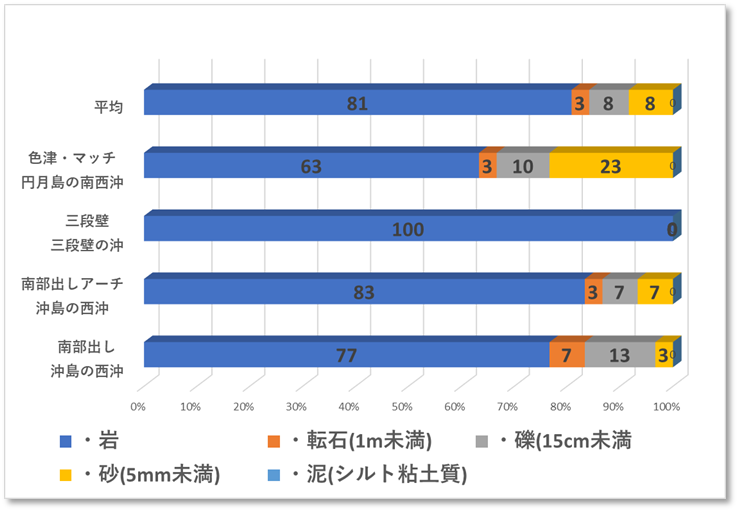

〇サンゴの生息状況 4地点の生サンゴ被度は1%程度であり、白化したサンゴや1年以内にへい死したサンゴはみつからなかった。サンゴ優占型形状は塊状のキクメイシ類、それ以外はトゲトサカ等のソフトコーラルであった。円月島の南西に位置する色津・マッチ(ダイビングポイント名)でミドリイシ属の稚サンゴ1個体(長径8cm)を確認した。
〇色津・マッチは、白浜のランドマーク円月島の南側の沖に位置し、水深は5mから15m、水底は平坦な砂地から岩礁が立ち並び、水路やアーチ・穴が多く富む複雑な地形が続く。底質は岩礁・砂地・礫・転石が混じる多様性に富む地形であった。
〇三段壁は、白浜の代表的な観光地である三段壁の沖に位置し、水深は5mから25m、水底からそそり立つ巨岩や整然と区切られたような階段状の構造は、人工的な遺跡のようにも見える特徴的な地形が広がっていた。
〇南部出しは、天神崎から南西に約3kmの地点に位置し、水深は5mから15m、岩礁の周囲は平坦な砂地(水深18m)に囲まれ、巨岩がそそり立ち、クレパスのような割れ目、オーバーハング、アーチなど地形は複雑であった。底質は岩礁・巨岩・転石・礫・砂地と多様性に富む地形であった。
〇南部出しアーチは、上記の南部出しから北東へ50m程に位置し、水深は5mから15m、岩礁の周囲は平坦な砂地(水深18m)に囲まれ、東西に伸びる岩礁には、アーチ・トンネル・オーバーハングと地形は複雑であった。底質は岩礁・巨岩・転石・礫・砂地と多様性に富む地形であった。
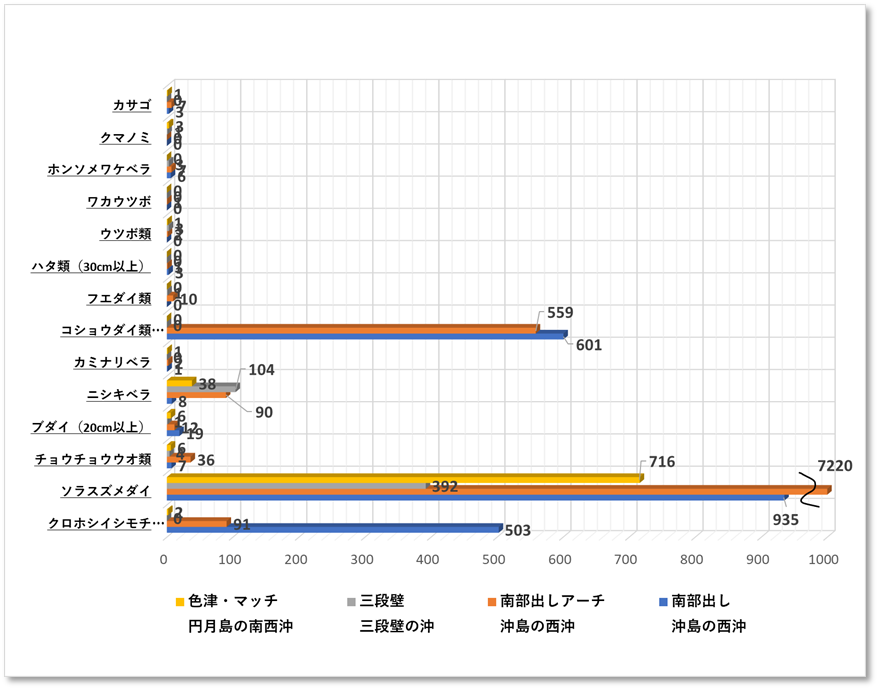
〇魚類 最も多様性に富む地点 南部出し・南部出しアーチは魚類の種類と数が多く、他地点よりも多様性に富んでいた。調査対象種以外ではキンギャハナダイやキンメモドキ数千匹の群、タカベ・アジ・スズメダイ・イワシの群れ、それら小魚を捕食しようとする大型のハタ類であるオオモンハタやクエ、回遊魚のカンパチなどがみられた。
〇無脊椎動物 ガンガゼ類がもっとも多くみつかった。調査対象種以外ではアオリイカ十数匹の小集団や岩肌を一面に覆うタワシウニを確認した。
🔲令和2年(2020年)被害状況調査

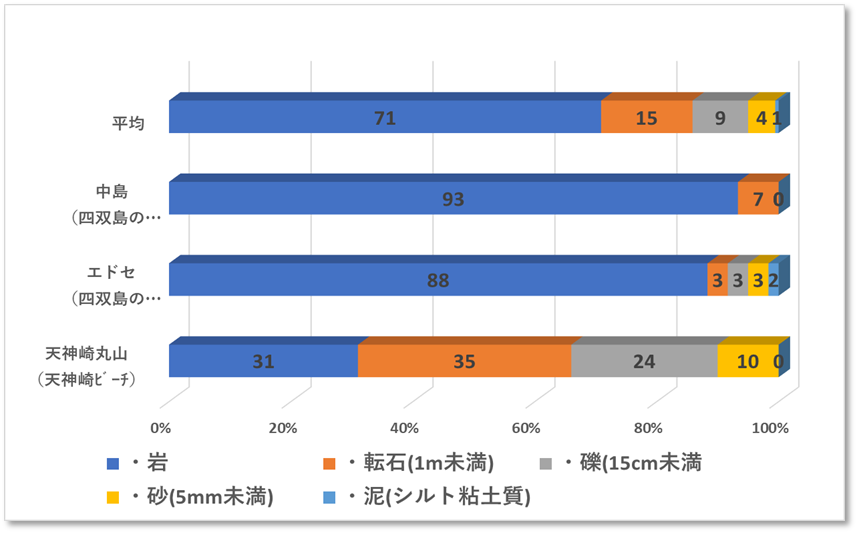

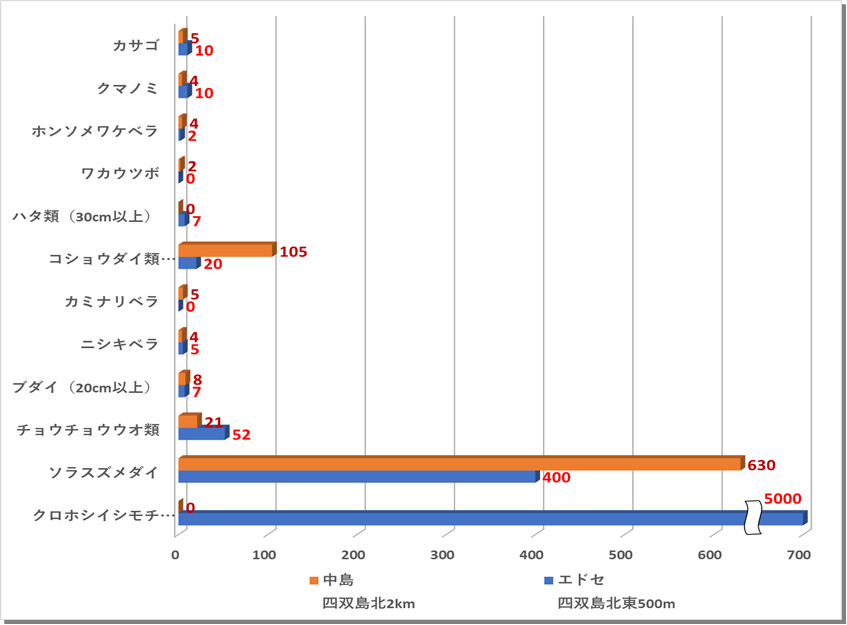
〇サンゴの生息状況
田辺市天神崎丸山の生サンゴ被度は32%、サンゴ優占型形状は樹枝状のエダミドリイシサンゴであった。稚サンゴは1㎡あたり3個であった。また、死サンゴ被度は1.3%であった。
白浜町中島の生サンゴ被度は1%程度であった。白浜町エドセの被度は1%未満であり、2地点において白化したサンゴや1年以内にへい死したサンゴはみつからなかった。2地点のサンゴ優占型形状は塊状のキクメイシ類、それ以外はトゲトサカなどのソフトコーラルであり、ミドリイシ属の稚サンゴは確認できなった。
特に中島ではウミトサカ類が多く、垂直面の岩肌を覆うように密集し生息していた。
〇中島は、白浜の番所鼻灯台から北へ2km弱に位置し、水深は5mから30m、岩礁の水深8mから観測塔が設置され、水温や風速など計測している。底質は岩礁が主であり、整地のための転石が集まるところもあった。
〇エドセは、白浜の番所鼻灯台から北西へ1km弱に位置し、水深は5mから20m、岩礁には幅8m奥行き5m程のカマボコ状の大きな穴があった。底質は岩礁・巨岩・転石・礫・砂地と多様性に富む地形であった。
〇天神崎丸山は、日本のナショナルトラストの先駆けの地であり、干潮時には平たい岩礁(海食台)が現れる。水深はタイドプールの0mから7m、底質は転石・礫・砂で構成されていた。
〇魚類 エドセにはクロホシイシモチが多く生息し5000匹と吐出していた。ついで多いのはソラスズメダイであり、中島では630匹、エドセでは400匹であった。次に、中島のコショウダイ類(イサキ)63匹、エドセのチョウチョウウオ類37匹であった。昨年度のクマノミの平均値は、1匹で数センチの小型の個体であったが、本年度は7匹、中には成魚となりペアで卵を育てる様子を確認した。
〇無脊椎動物類 ガンガゼ類がもっとも多くみつかった。
🔲令和3年(2021年)被害状況調査

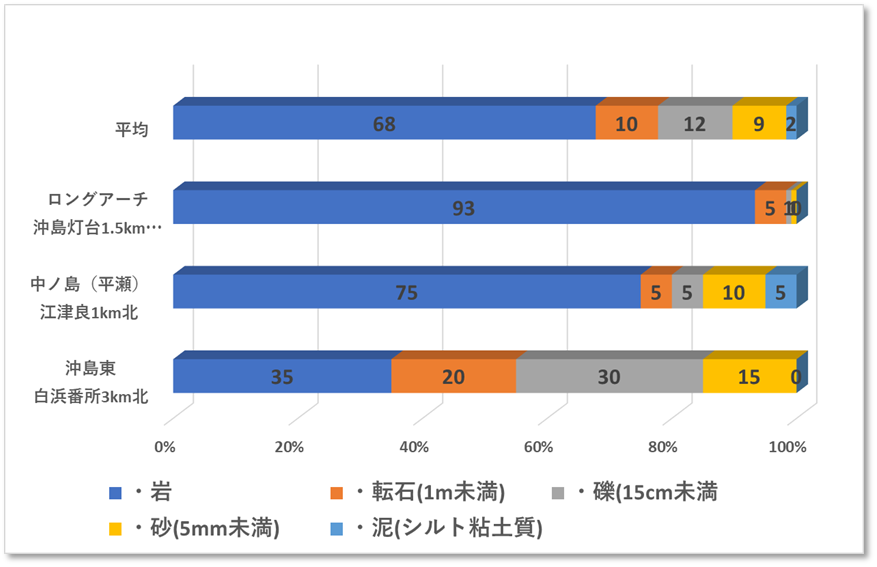

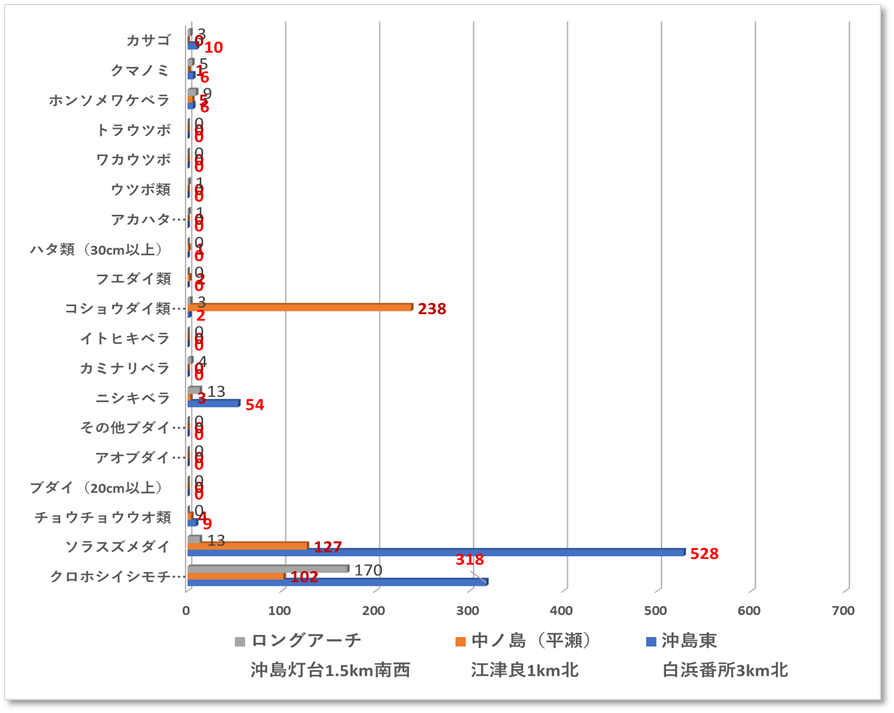
〇サンゴの生息状況
ロングアーチの生サンゴ被度は1~5%、優占型は塊状のキクメイシ類であった。また、ミドリイシ属の稚サンゴは合計で8個確認し、その内訳は5~10cm未満のサイズが1個、10~15cm未満が4個、15~20cm未満が2個、20cm以上が1個であった。
沖島東の生サンゴ被度は1%未満であった。また、稚サンゴは5~10cm未満を1個確認した。
中ノ島の生サンゴ被度は1%未満であった。
上記の3地点で、白化したサンゴや1年以内にへい死したサンゴはみつからなかった。特に中ノ島では、トゲトサカなどのウミトサカ類やウミウチワ・ヤギ類などソフトコーラルの種類と数を多く確認した。
〇沖島東側の地先であり、0mから10m、底質は多様性に富むが、水深は浅く、平坦な地形であった。
〇ロングアーチは沖島灯台から南西に600m弱に位置し、水深は5mから25m、岩礁は台形状であり周囲は砂地と岩礁が広がっていた。岩礁の中心には全長20m程の長い海中のトンネルがあった。底質は岩礁・砂地・礫・転石が混じる多様性に富む地形であった。
〇中ノ島は江津良浜から北へ1km強に位置し、水深は5mから23m、直立する岩礁の最浅部は5mの棚状の地形が連続し、岩礁には多くの亀裂が見られ、オーバーハング・アーチが連続するなど変化に富んでいた。底質は岩礁・砂地・転石・礫・泥が混じる多様性に富む地形であった。
🔲令和元年度から令和3年度までの3年間の本調査のまとめ・考察
3.サンゴ群集浅海域調査まとめ・考察
令和元年度から令和3年度までの3年間の本調査のまとめと、独自調査の知見も引用し、浅海域の生物多様性調査である被害状況調査、および重点海域調査について、以下に考察を述べる。
(1)世界北限域の高被度サンゴ群集消失
2017年迄、田辺湾沖島周辺や白浜町沿岸の円月島、四双島、権現崎は、卓上型(テーブル状)のミドリイシ属サンゴが広域にわたり高被度に生息する世界北限域の大サンゴ群集であった。
2018年1月下旬、田辺市・白浜町のダイビング事業者からサンゴ白化の緊急報告が寄せられた。要約すると、「白化は5割以上に達している」、「白化死したサンゴは1割以上」という。
「平成29年度サンゴ群集モニタリング調査及びオニヒトデ等調査駆除の結果及び報告会」(環境省2018.4)によると、天神崎ビーチ(天神崎丸山)では2017年12月20日時点で、サンゴ被度32%・白化率5%であったが、2018年2月27日時点では被度25%・白化率90%(うち死亡率20%)に減少した。水温は最低10.5℃を記録し、調査時水温は13.7℃でった。
沖島コーラルガーデン(ダイビングポイント名で沖島の北側)では、2017年12月22日時点で、被度72%・白化率5%以下であったが、2018年2月27日には、被度10%・白化率95%(うち死亡率85%)・調査時水温14℃であった。
ニシザキサンゴの真ん中(ダイビングポイント名で、沖島西側のサンゴ密集地)では、2018年2月27日時点で、被度13%(白化前推定被度85%)・白化率95%(うち死亡率85%)・調査時水温14.6℃であった。
四双島南では、2018年3月13日時点で、被度5%(白化前推定被度50%)・白化率100%(うち死亡率95%)・調査時水温13.4℃であった。
権現崎では、被度5%(白化前推定被度50%)・白化率100%(うち死亡率95%)・調査時水温13℃と報告された。
一方、2018年は12年ぶりの黒潮大蛇行の年に当たり、且つ、1月・2月の西日本の平均気温平年差は-1.2℃を記録(気象庁「平成30年冬の天候の特徴とその要因について」)した。これは1986年冬以降32年ぶりの最も寒い冬、いわゆる「大寒波」となった。
これらの要因により、田辺白浜海域(以降、「本海域」と略す。)の水温は、1月中旬から3月中旬まで2か月間、13~14℃台(時には10℃台)を継続した。サンゴの耐低水温限界とされる15℃以下を2か月の長期にわたり下回った。
串本海域の調査報告(御前1984,1985)によると、14℃ 台の水温が1か月以上続くとサンゴに白化やへい死が生じ、13℃台の期間が20日以上続くとサンゴの大量へい死に至ると推察されていることより、本海域のサンゴは低水温の長期化により、同時一斉に大量へい死し、壊滅的な被害(以降、「2018年の低水温ショック」と略す。)になった。
(2)サンゴ群集の遷移
① 沖島(沖島西と北)
2004年まで沖島西と沖島北のサンゴ被度は80%と高被度であったが、2004年7月から2005年8月の1年2か月の黒潮大蛇行の影響により低水温が長引き、38%に減少した。その後、回復し60%前後を維持したが、2018年の低水温ショックにより、壊滅的に減少し1%程度になった。以降、サンゴ被度は1%程度を継続しており、明確なサンゴ被度の増減は認められない。
② 四双島
2008年まで四双島のサンゴ被度は65%と高被度であったが、2009年のオニヒトデ大発生による食害により、サンゴは急激に減少推移し、2018年の低水温ショックにより壊滅的に減少し1%程度となり、本調査においても明確なサンゴ被度の増減は認められない。
③ 円月島
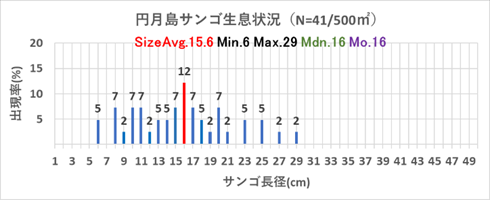
2008年まで円月島のサンゴ被度は70%と高被度であったが、2009年のオニヒトデ大発生による食害により、サンゴは急激に減少推移し、2018年の低水温ショックにより壊滅的に減少し1%程度となり、本調査においても明確なサンゴ被度の増減は認められない。
本海域の主要サンゴ群集被度の経年変化を以下の図8.1に示す。
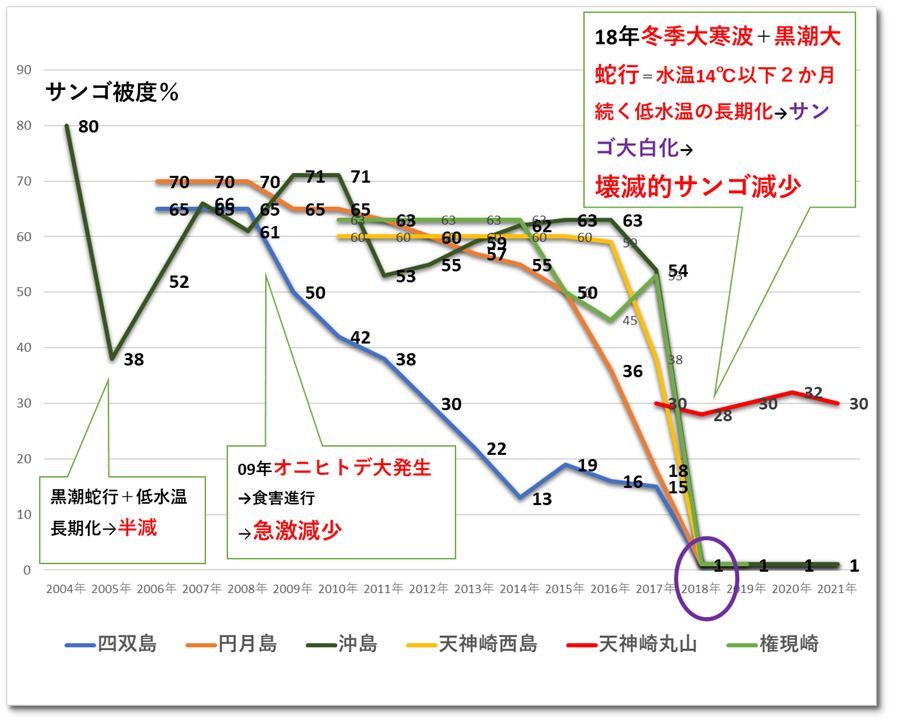
※本調査および独自調査より作図。
図Ⅲ.8.1 サンゴ生息状況の遷移
(3)サンゴ群集の再生状況
サンゴの新規加入量の多寡は、サンゴ群集の回復に密接に関係すると考えられ、群集回復の予測には重要な指標の一つである。
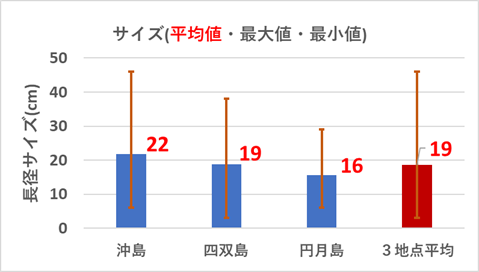
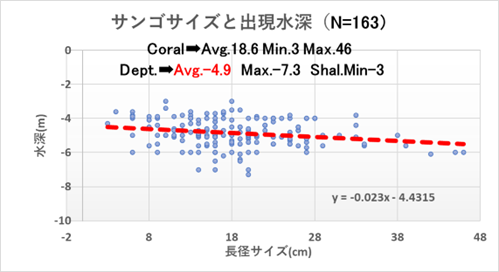
重点海域のモニタリング地点である沖島西と北・四双島・円月島のサンゴ被度の増減は明確ではなかったが、サンゴ再生に大きく寄与する成長の早いミドリイシ属の稚サンゴの増加が顕著であったので、以下図9.1から図9.4に、集約データを表12.1から12.4に、状況写真を別添の「重点海域のミドリイシ属サンゴ再生調査の状況写真」に示す。
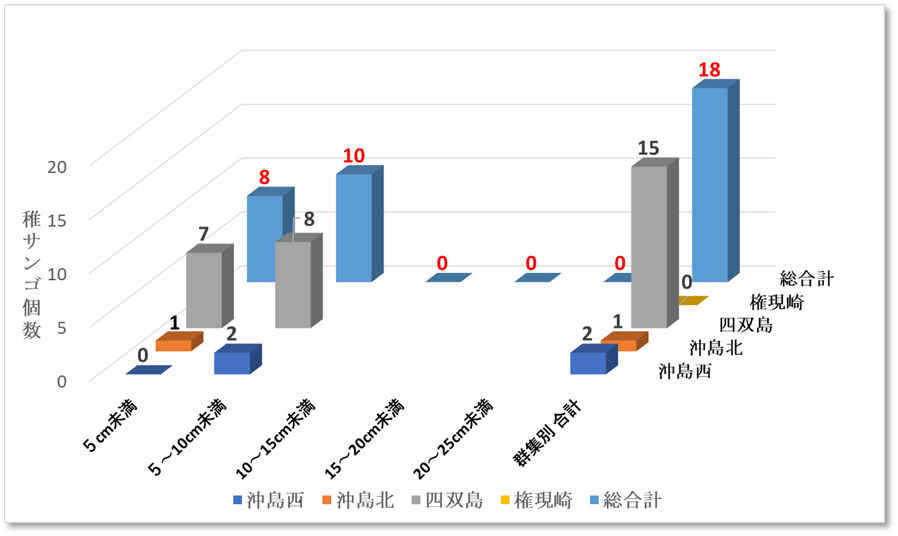
図Ⅲ.9.1 2019年ミドリイシ属稚サンゴサイズ別集計図
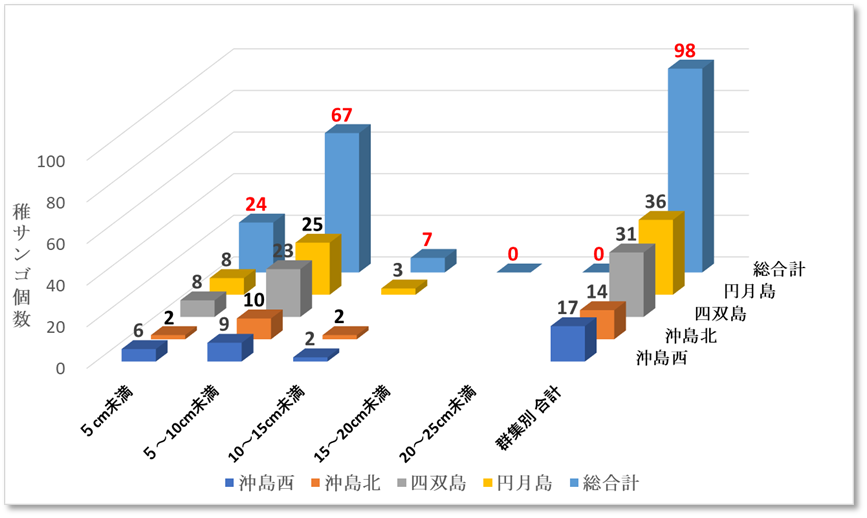
図Ⅲ.9.2 2020年ミドリイシ属稚サンゴサイズ別集計図
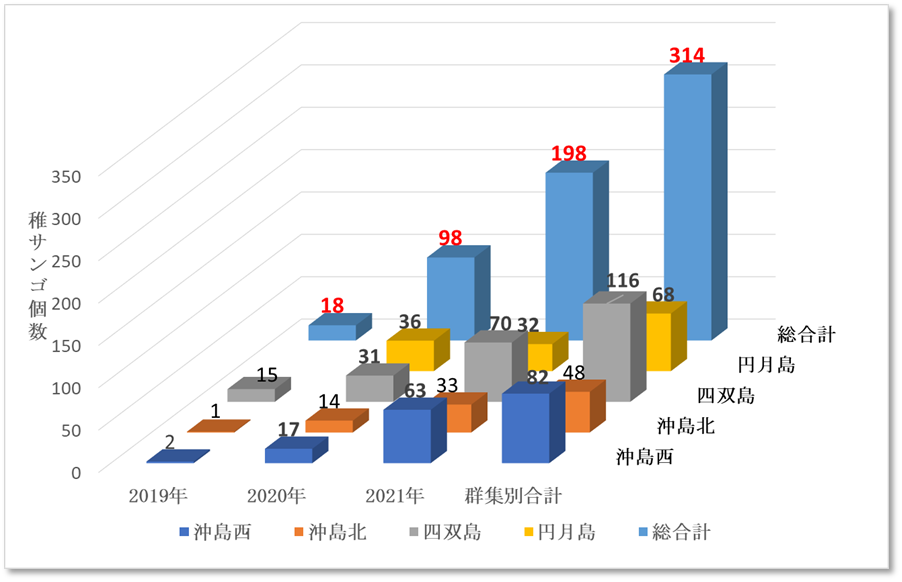
図Ⅲ.9.4 ミドリイシ属稚サンゴ年度別集計図
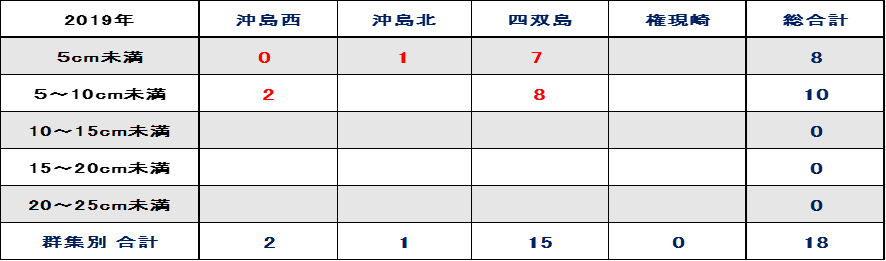
表Ⅲ.12.1 2019年ミドリイシ属稚サンゴサイズ別集計
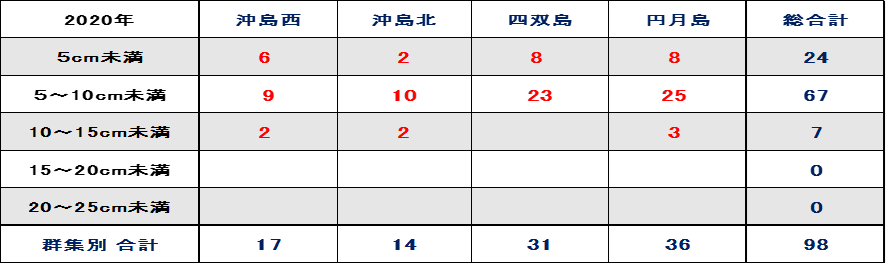
表Ⅲ.12.2 2020年ミドリイシ属稚サンゴサイズ別集計
表Ⅲ.12.3 2021年ミドリイシ属稚サンゴサイズ別集計
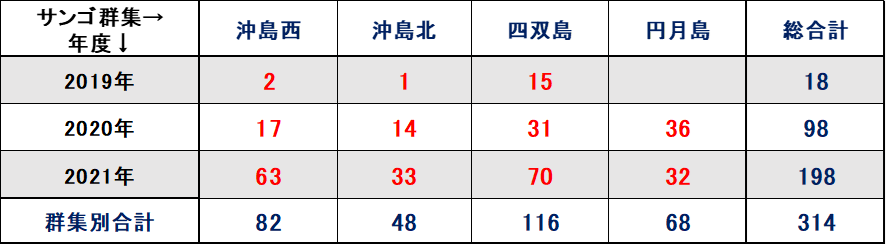
表Ⅲ.12.4 ミドリイシ属稚サンゴ年度別集計図
➢2019年ミドリイシ属稚サンゴ出現
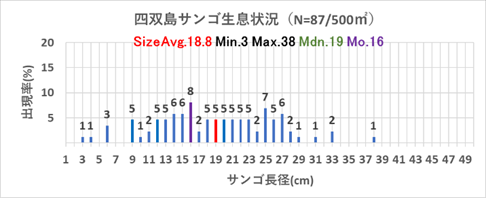
2019年の稚サンゴの加入について、表12.1、図9.1をみると、四双島で5cm未満の稚サンゴが7個、5~10cm未満が8個、沖島西では5~10cm未満が2個、沖島北では5cm未満1個、2019年の稚サンゴ総合計は18個であった。
予想以上に早い稚サンゴの出現に、新規に加入した稚サンゴなのか、2018年の低水温ショックをのりこえ生残した個体なのか、評価が難しかった。
➢2020年ミドリイシ属稚サンゴ増加
2018年の低水温ショックの年に浅い水深帯では見つけられなかったミドリイシ属の稚サンゴは、2019年から数メートルの浅い水深で発見されるようになった。水深2から4mのミドリイシ属のサンゴを主に調査を実施したところ図9.2、表12.2に示す通り、総合計で98個に増加した。
これら稚サンゴは、1㎡あたり最も多いところで5群体であったが、成長し群体が融合(合体)した形跡も多く、合体前は8個程あったと推察する。それでも回復傾向であるとする10群体/㎡には満たなかった。しかし、2018年時点で浅い水深で見つけられなかったミドリイシ属の稚サンゴが予想以上に発見できたことに再生の兆しを感じる。
➢稚サンゴは新規加入と推測
2018年に浅い水深帯で発見されなかった(見落としも考えられる)ミドリイシ属の稚サンゴが2020年に多く発見されたことは、2018年の低水温ショックを乗り越え生残した稚サンゴか、或いは、新たにサンゴ卵や幼生が他の南方の海域より流れ着き、着生し成長した新規加入サンゴかのどちらかである。
稚サンゴが2018年の低水温ショックを乗り越え生残することは、以下の観点より困難と考えられる。2020年の調査で多く見つかった数メートルの浅い水深帯は、それ以深の水深帯より水温が低い。さらに、干潮時ともなれば大寒波の寒風に直接さらされることもあり得る。このような状況で2018年の低水温ショックを稚サンゴだけが生き残ることは容易ではない。
一方、本海域で育つクシハダミドリイシサンゴやエンタクミドリイシサンゴの成長スピードは、経験則から年間に4から10cm程度と推測され、表12.2の5~10cm未満のサイズが67群体(68%)最も多く、1から2年目の成長したサイズとして妥当である。また、10cm以上のサイズは7群体(7%)であるが複数の稚サンゴが融合した形跡がある。以上により、稚サンゴは新規加入であると推測する。
2018年の低水温ショックではサンゴだけでなくクマノミも姿を消し、生残するクマノミを発見することが難しい状況にまで激減したが、2019年には数センチの幼魚のクマノミが確認されるようになり、2020年にはペアリングした成魚が卵を守る様子が確認できた。このことは、他海域から新たなクマノミの稚魚が流れ着き成長したと考えられる。クマノミの産卵時期はサンゴの産卵と大きくずれはなく、同様にサンゴ卵やその幼生も流れつき成長したと考えることが自然である。
➢2021年ミドリイシ属稚サンゴ順調に増加
表12.3、図9.3に示す通り、稚サンゴのサイズと数から評価すると、稚サンゴは大きく成長し、且つ新規加入の小さな稚サンゴも確認できる。なかには20cm以上の群体も確認でき、順調に育っていると考える。
➢サンゴ再生の兆しあり
表12.4、図9.4の示す通り、2019年にミドリイシ属の稚サンゴ18個が出現し、2020年に98個に増加、2021年には198個確認できた。2018年の低水温ショック直後は全く発見することができなかった稚サンゴが、3年あまりで総合計314個確認できたことは、明らかに稚サンゴが本海域に新規加入し、成長している証である。
➢サンゴは回復傾向
2021年調査では、稚サンゴの1㎡あたり最も多いところで10群体を確認した。回復傾向であるとする10群体/㎡を満足した結果となった。今後は低水温や高水温の長期化、超大型台風の波浪等による連続的かく乱、オニヒトデやレイシ貝類による食害圧が大きくない限り、本海域のサンゴ群集は順調に回復すると推測する。
ちなみに、2018年の低水温ショック以降、サンゴ群集の被度が明確に増加しないのは、現状のサンゴ類の優占種がキクメイシ類であり成長が遅いことの理由による。今後、成長の早いミドリイシ属の稚サンゴが優占種となれば、順調にサンゴ被度が増加すると推測する。
(4)浅海域調査地点の多様性評価
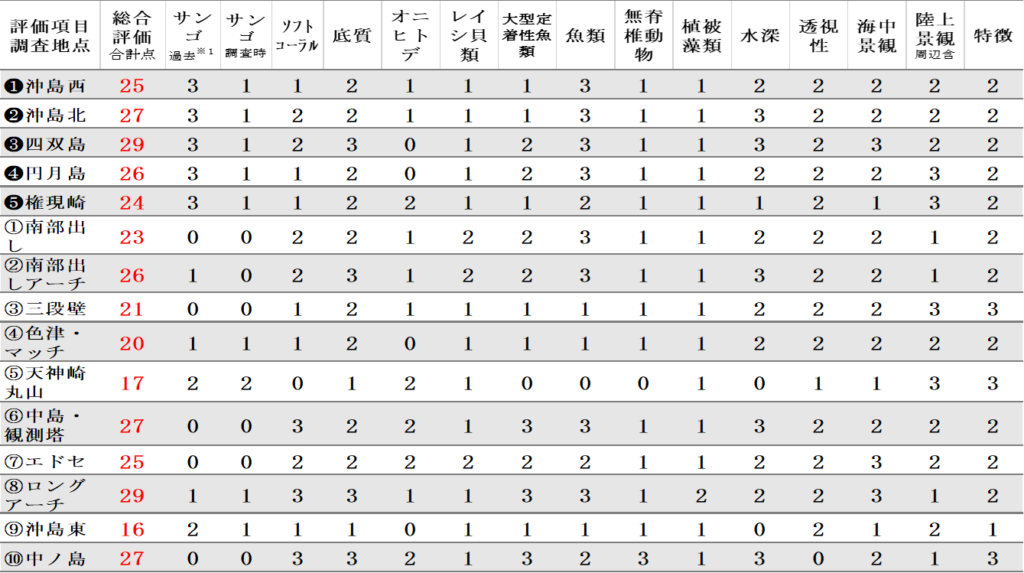
浅海域の各調査地点の多様性を以下の通り評価した。なお、評価項目と評価点を表13.1に、総合評価結果を表13.2に示す。
➢高評価地点
総合評価が最も高かったのは、29点の❸四双島と⑧ロングアーチであった。その次に高ったのは27点の❷沖島北、⑥中島(観測塔)、⑩中ノ島であった。
➢低評価地点
総合評価が最も低かったのは、16点の⑨沖島東、その次に低かったのは⑤天神崎丸山の17点であった。
表Ⅲ.13.1 評価項目と評価点

表Ⅲ.13.2 浅海域調査地点の多様性総合評価
4.サンゴ捕食生物調査まとめ・考察
令和元年度から令和3年度までの3年間の本調査のまとめと、独自調査の知見も引用し、サンゴ捕食生物の調査のまとめと考察を以下に述べる。なお、調査方法などの詳細については、別添の「レイシ貝類について(抜粋版説明用)」および「オニヒトデ駆除マニュアル(抜粋版説明用)」を参照のこと。
4.1.天神崎丸山のレイシ貝類調査まとめ・考察
2020年度の調査結果より、天神崎丸山の死サンゴの主要因はレイシ貝類による食害死と推測された。コドラート枠内のレイシ貝類は1㎡あたりに換算すると5個程であり多くはなさそうであるが、均一に分散はせず密集する場所では食害によりへい死したエダミドリイシサンゴが点在していた。
また、食害が集中する場所のレイシ貝を調べると殆どがクチベニレイシガイダマシであった。過年度は同種に加えシロレイシガイダマシやヒメシロレイシガイダマシも多く確認した。
これらレイシ貝類は大発生しオニヒトデのような深刻な被害を及ぼすことがあり、大集団が形成され始めるとサンゴの急激な被度低下が懸念されるので、調査の都度、恣意的な除去を細々と実施していた。
(1)レイシ貝類の生息状況の評価規準の設定
レイシ貝類の生息状況を判断するため、過去の調査や除去の経験値より、暫定的に以下の通り評価規準を設定した。この数値は1人1回、30分から45分程度の潜水によるレイシ貝類の除去数を想定している。また、本活動にあたる調査員は、除去調査に慣れていることを前提とする。
表Ⅲ.14.1 レイシ貝類生息状況評価規準
| レイシ貝類の除去数(1人1回) | 評価/現象 | 保全指針 |
| ①100個未満 | 自然発生の生息数/サンゴ1群体(または1㎡)に数個~十数個 | 自然環境に任せ見守るが、50個以上は要注意 |
| ② 100個~500個未満 | 要警戒/サンゴ1群体(または1㎡)に数十個以上の集団顕著化、サンゴ被度低下傾向 | 計画的な除去、モニタリング継続が必要 |
| ③ 500個~1000個未満 | 大発生/サンゴ白化と密集団多見、サンゴ被度顕著に低下、景観荒廃進む | 保全可能な範囲を設定、計画的に徹底した除去を継続 |
| ④ 1000個以上 | 絶滅危惧/サンゴ群体上に砂利を敷き詰めたように密集、いたる所でサンゴ白化と被度低下、景観著しく荒廃 | 最低限の保全範囲を決め、徹底した除去を繰り返し、保全範囲外のレイシ貝の収束を待つ |
※慣れている人と初めての人の除去比は、2:1~5:1とする。
(2)天神崎丸山のレイシ貝類の生息状況
2021年度の調査除去の結果より、レイシ貝類1人・1回あたりの除去数は、78個であった。また、1㎡あたりの生息数は5個であり、表14.1の評価規準にてらし判断すると、自然発生の生息数であり、レイシ貝類の食害により、サンゴが急激に減少するような状況となる生息数ではなかった。
ただし、レイシ貝類は均一に分散せず、密集する場所では、食害によりへい死したエダミドリイシサンゴの群体が点在していた。
また、本年度は除去範囲が狭く、エダミドリイシサンゴ密集域に限った除去なので評価の精度は低い。
よって、今後は除去範囲を拡大し再度の除去活動を実施した後に、最終的な生息状況を評価し、保全の是非を判断する必要があると考える。
4.2.オニヒトデ調査まとめ考察
2018年の低水温ショックにより一度は全滅し、以降は生残や新規加入するオニヒトデは発見されなかったが、2020年10月に田辺市のダイビング事業者により、田辺湾沖島の西側500m地点の水深15mに、体長20cm程のオニヒトデ1個体が発見された。
しかし、本年度の調査ではオニヒトデは発見されず、また、現地ダイビング事業者のヒヤリングにおいてもオニヒトデは発見されていない。
亜熱帯性の魚類であるクマノミは、2018年の低水温ショックにより、一時期はその姿を消していたが、2019年には数cmの稚魚が確認され、2020年からは卵を育てる様子が多く確認されている。また、本調査海域において、ミドリイシ属の稚サンゴも新規加入が確認できた。これらは、南方海域から潮流や台風に運ばれ、新規に加入した個体であると考えられ、同様にオニヒトデの卵や幼生も本海域に流れ着いた個体が定着する可能性がある。産卵可能な成体にまで育つ期間は3年から4年であり、2019年から順調に幼生が流れ着いていたとすれば、まさに本年が危惧される年であったが、1個体も確認はできなかった。今後も注意深く調査を継続する必要がある。オニヒトデの生息状況の経年変化を以下の図10に示す。
4.2.オニヒトデ調査まとめ考察
2018年の低水温ショックにより一度は全滅し、以降は生残や新規加入するオニヒトデは発見されなかったが、2020年10月に田辺市のダイビング事業者により、田辺湾沖島の西側500m地点の水深15mに、体長20cm程のオニヒトデ1個体が発見された。
しかし、本年度の調査ではオニヒトデは発見されず、また、現地ダイビング事業者のヒヤリングにおいてもオニヒトデは発見されていない。
亜熱帯性の魚類であるクマノミは、2018年の低水温ショックにより、一時期はその姿を消していたが、2019年には数cmの稚魚が確認され、2020年からは卵を育てる様子が多く確認されている。また、本調査海域において、ミドリイシ属の稚サンゴも新規加入が確認できた。これらは、南方海域から潮流や台風に運ばれ、新規に加入した個体であると考えられ、同様にオニヒトデの卵や幼生も本海域に流れ着いた個体が定着する可能性がある。産卵可能な成体にまで育つ期間は3年から4年であり、2019年から順調に幼生が流れ着いていたとすれば、まさに本年が危惧される年であったが、1個体も確認はできなかった。今後も注意深く調査を継続する必要がある。オニヒトデの生息状況の経年変化を以下の図10に示す。
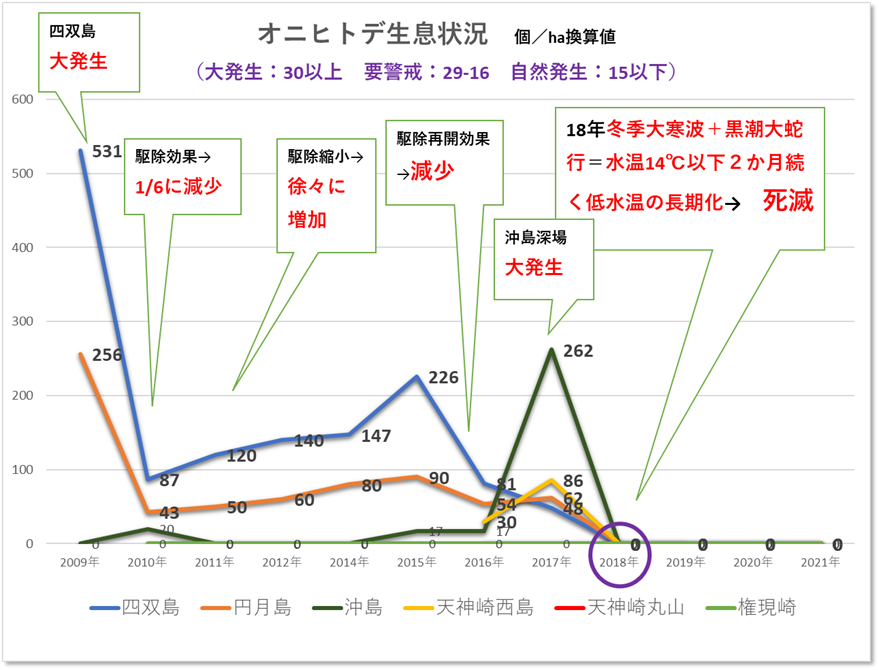
図Ⅲ.10 オニヒトデ生息状況経年変化
🔲調査等の状況写真









実施済み調査
●2021年度生物多様性調査・被害状況調査・深海域調査
●2020年度生物多様性調査・被害状況調査・深海域調査
●2019年度生物多様性調査・被害状況調査・深海域調査
●2018年度権現崎リーフチェック
●2018年度沖島リーフチェック
●2018年度円月島リーフチェック
●2017年度権現崎リーフチェック
●2017年度沖島リーフチェック
●2017年度円月島リーフチェック
●2017年度権現崎リーフチェック
●2016年5月29日第5回四双島リーフチェック
●2016年5月28日第13回沖島リーフチェック
●2015年6月7日第12回沖島リーフチェック
●2014年6月8日第11回沖島リーフチェック
●2014年6月7日第4回四双島リーフチェック
●2011年11月27日第2回四双島リーフチェック
●2011年11月26日第9回沖島リーフチェック
●2012年11月4日第3回四双島リーフチェック
●2012年11月3日第10回沖島リーフチェック
●2010年5月30日第8回沖島リーフチェック
●2010年5月29日第1回四双島リーフチェック
●2009年5月31日第7回沖島リーフチェック
●2008年11月16日第6回沖島リーフチェック
●2007年5月27日第5回沖島リーフチェック
●2006年5月27日、28日、7月2日第3回、4回沖島リーフチェック
●2005年5月28日、29日第2回沖島リーフチェック
●2004年5月29日、30日第1回沖島リーフチェック
報告・感想・コメント

【2015-16年リーフチェック(サンゴ生物調査報告】
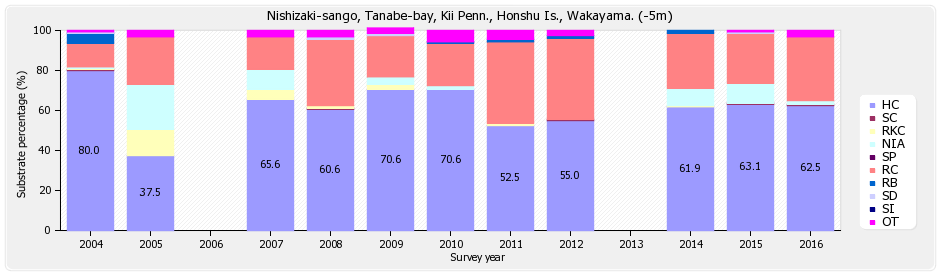
沖島
底質調査の浅い(-5m)調査ラインは、ハードコーラル(HC)62.5%、ソフトコーラル(SC)0.6%、最近死んだサンゴ(RKC)0%、富栄養化の指標となる海藻(NIA)1.9%、海綿類(SP)0%、岩(RC)31.9%、礫(RB)0%、砂(SD)0%、シルト(SD)0%、その他(OT)3.1%となっている。
※図B-1①沖島 底質分類(浅)の経年変化 を参照下さい。
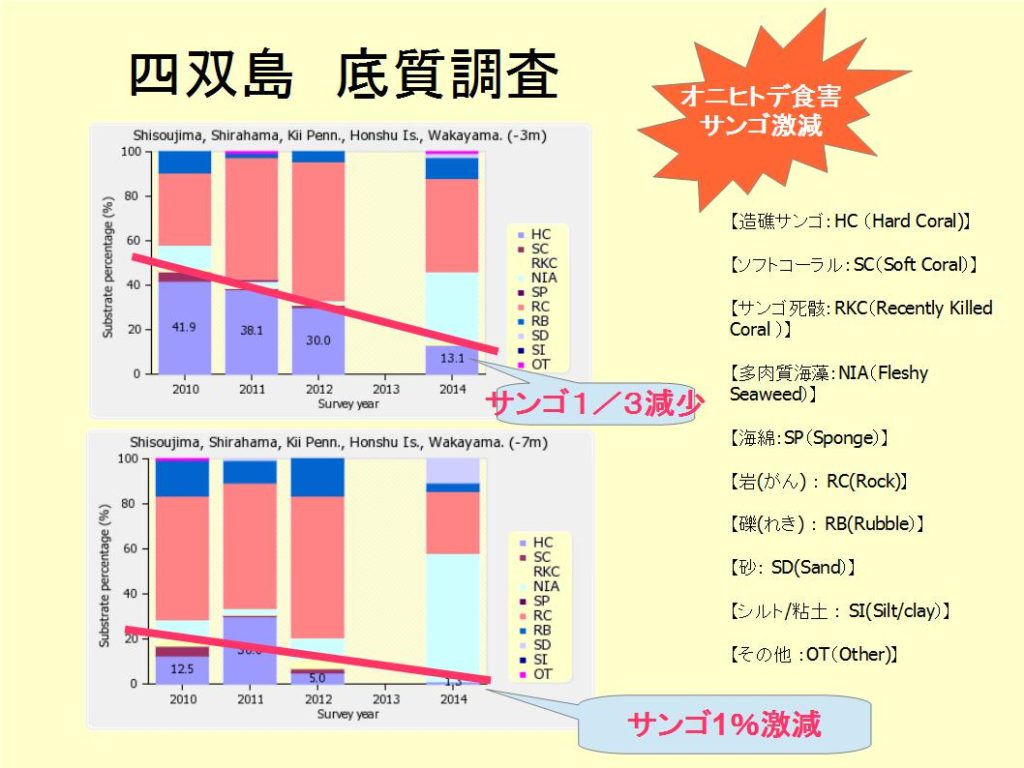
経年変化をみると、2004年のハードコーラル(HC)は80%から、2005年の37.5%と大きく減少した。これは2005年の低水温(真夏でも-5mで18℃)の影響、及び調査ラインの一時的なずれによるデータのバラツキの結果と考えられるが、その後、回復し6割以上を保っている。
底質調査の深い(-10m)ラインは、ハードコーラル(HC)28.7%、ソフトコーラル(SC)0.6%、最近死んだサンゴ(RKC)1.3%、富栄養化の指標となる海藻(NIA)54.4%、海綿類(SP)0%、岩(RC)11.3%、礫(RB)0.6%、砂(SD)3.1%、シルト(SD)0%、その他(OT)0%となっている。
※図B-1②沖島 底質分類(深)の経年変化 を参照下さい。
経年変化をみると、2016年のハードコーラル(HC)が過去最低の28.7%となったが、海藻(NIA)が54.4%とサンゴ面に覆いつくしている影響でサンゴの被覆度が小さくなっていると考えられる。 本年は他の海域(例えば、円月島、四双島、海岸に近い田尻)などにおいても海藻は多い傾向にあった。 尚、魚類、無脊椎動物についてのデータは割愛する。


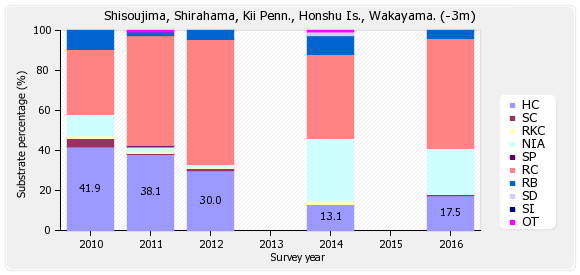
四双島
底質調査の浅い(-3m)調査ラインは、ハードコーラル(HC)17.5%、ソフトコーラル(SC)0.6%、最近死んだサンゴ(RKC)0%、富栄養化の指標となる海藻(NIA)23.1%、海綿類(SP)0%、岩(RC)55%、礫(RB)3.8%、砂(SD)0%、シルト(SD)0%、その他(OT)0%となっている。
※図B-2① 四双島 底質分類(浅)の経年変化 を参照下さい。
経年変化をみると、2010年のハードコーラル(HC)は41.9%から、2016年の17.5%と半減した。これはオニヒトデによる食害が原因である。
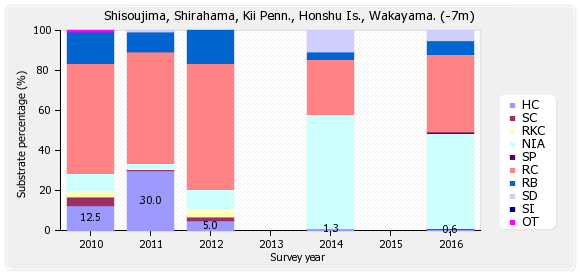
底質調査の深い(-7m)ラインは、ハードコーラル(HC)0.6%、ソフトコーラル(SC)0.6%、最近死んだサンゴ(RKC)0%、富栄養化の指標となる海藻(NIA)47.5%、海綿類(SP)0.6%、岩(RC)38.8%、礫(RB)6.9%、砂(SD)5%、シルト(SD)0%、その他(OT)0%となっている。
※図B-2②四双島 底質分類(深)の経年変化 を参照下さい。
経年変化をみると、2010年のハードコーラル(HC)は12.5%であったが、2014年には1.3%に減少し、2016年は0.6%と減少した。これはオニヒトデによる食害が原因である。
2011年のハードコーラル38.1%は、調査ラインが浅い側(サンゴご多い方向)にズレたための結果で、サンゴが回復した訳ではない。 尚、魚類、無脊椎動物についてのデータは割愛する。
2014年までの纏め
オニヒトデの食害により
「四双島」に生息するサンゴは激減しました。
対応策を講じないまま放置した場合、
数年後には壊滅状態になると推測します。
沖ノ島に生息するサンゴは、2014年6月時点では、
オニヒトデ等による目だった食害は確認できません。

TSコメント【調査ラインS】 HCの割合は、一昨年の 30%から大きく減り、13%であった。 一方、NIAが、一昨年の1.3%から大きく増えて、31%であった。 HCの減少は、主にオニヒトデによる食害とみられ、 調査ライン後半の一部分を除いて、食べつくされたに近い状態と なっている。 オニヒトデは、調査範囲内では1個体しか見つからず、 一昨年の7個体より減少しているが、 さらに浅い海域に10個体程度見られ、浅い方に移動していって いるとみられる。 大きさはいずれも、25~30cm大であった。 増加していた海藻は、主にヒロメであった。 海藻増加の要因としては、HCの減少によって繁殖可能な場所が増えたことの他に、 一昨年は調査を秋に行ったが、今年は春に行ったことも 影響していると思われる。 海藻の上には、シルトの堆積が多少みられるが、 岩の上にはシルトはほとんどみられず、岩の上の砂が若干多いように見える。 魚類・無脊椎動物では、海藻が多い割には、ガンガゼ類が1個体のみ (一昨年は10個体)と少なかった他は、大きな変化は見られなかった。 【調査ラインM】 昨年は調査が行えなかったため、2年ぶりの調査であった。 HCの割合は、一昨年の 5%からさらに減少し、1%であった。 一方、NIAが、一昨年の 9.4%から大幅に増加し、57%であった。 HCの減少は、主にオニヒトデによる食害とみられ、 ほぼオニヒトデに食べつくされた感じである。 すでにこの周辺には、オニヒトデも見られない。 時期の問題もあるかもしれないが、海藻多い。 増加していた海藻は、主にヒロメであった。 海藻増加の要因としては、HCの減少によって繁殖可能な場所が増えたことの他に、 一昨年は調査を秋に行ったが、今年は春に行ったことも 影響していると思われる。 海藻の上には、シルトの堆積が多少みられるが、 岩の上にはシルトはほとんどみられず、岩の上の砂が若干多いように見える。 魚類、無脊椎動物については、オニヒトデが見られなくなったことの他は、 大きな変化は見られない。






















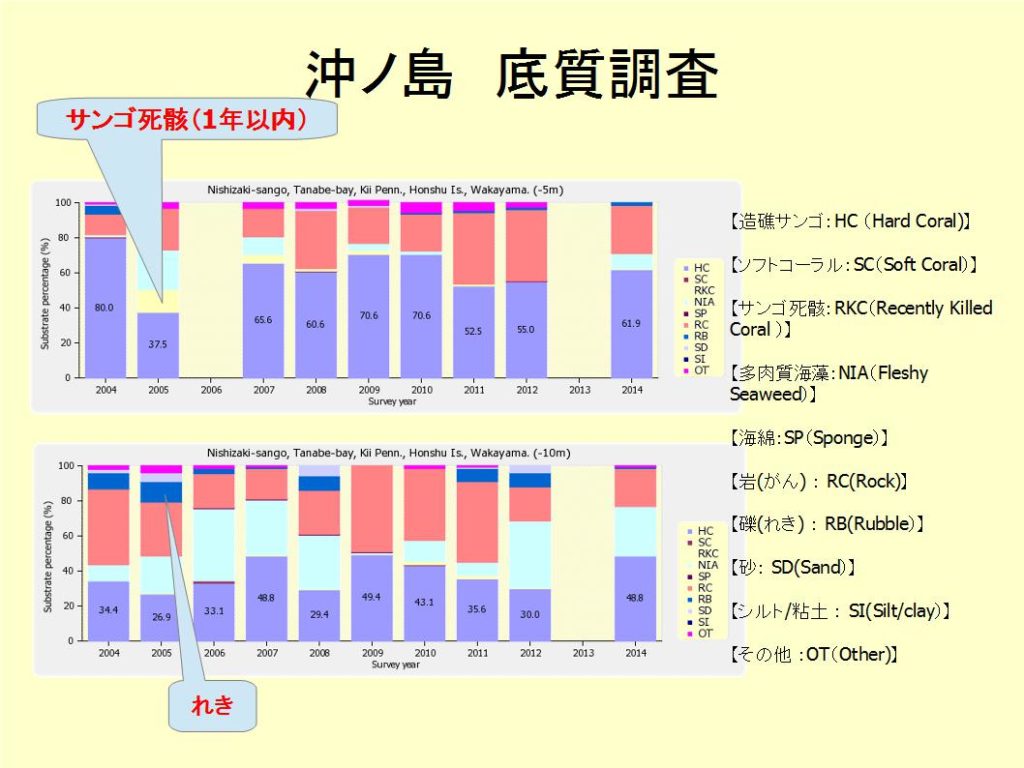
第9回 沖ノ島(ニシザキサンゴ) リーフチェック報告

第9回 沖ノ島(ニシザキサンゴ) リーフチェック報告
リーフチェック田辺2011 TSコメント
[-5m]
-5mの調査ラインは、田辺湾の中ほどにある島から続く斜面の、
傾斜がゆるくなっている部分の上で、テーブル状のエンタクミドリイシ、被覆状のニホンミドリイシが優占している場所に設置されている。
HCの割合は、昨年の70.6%から、52.5%に大幅に下落している。
これは、9月にごく近くを通過した、台風12号 “TALAS” および台風15号 “ROKE”の影響と思われる。
2度の台風により、テーブル状のサンゴが根こそぎ持ち去られたようで、サンゴがなくなり、岩盤が露出している所がめだち、大きく景観が変わってし まっていた。
また、テーブル状のエンタクミドリイシで、部分的な死亡が多く見られたが、サンゴ食巻貝による食害と思われる。
昨年の冬期に、水温11℃の冷水塊が長期間留まっていたために、
サンゴ食巻貝が大分減ったとのことではあるが、まだ影響が大きいようであり、今後も注意が必要である。
HCの割合の低下には、冬場の低水温の影響もあったかもしれない。
テーブル状サンゴ、被覆状サンゴともに、粘液をさかんに出しているコロニーがたくさん見られた。
9月の台風による大雨により、田辺湾に注ぎ込む河川の上流域で、
大規模な土砂崩れが発生したため、大量の泥が海に流れ出しており、台風から2ヶ月経過した調査時には、見た目では、生きたサンゴの上にシルトの堆積があるようには見えなかったが、シルトの影響があるかもしれない。
実際、調査地の沖側の水深15m付近の海底に堆積しているサンゴの破片の上には、シルトの堆積が見られる。
今後も長期にわたって雨が降った際に泥の流出が続く可能性があり、継続的な状況の監視が必要であると思われる。
魚類・無脊椎動物では、例年実施している春の時期に低気圧が来て開催できず、秋に調査を実施した影響で、小さなチョウチョウウオ類が多く見られた。
また、イサキの群れが付近を回遊していたため、これまでに見られない数が観測された。

[-10m]
-10mの調査ラインは、-5mの調査ラインに平行した、斜面の中ほどに設置されており、-15m付近からは砂地となっている。
この付近では、ニホンミドリイシはあまり多くなく、エンタクミドリイシの他、キクメイシなどいろいろな種類の被覆状、塊状のサンゴが見られる。
HCの割合は、昨年の43.1%から35.6%に下落しているが、-5mよりは下落幅が小さかったのは、被覆状・塊状のサンゴが多いため、-5mほど台風の影響を受けなかったためと思われる。
調査ラインのすぐ沖側には、破片化したサンゴが多数沈んでおり、台風によって破壊されたサンゴであると思われる。
-5mと同様に、部分的に死亡しているサンゴや、粘液を多く出しているサンゴも多く見られた。
魚類・無脊椎動物については、ガンガゼ類が昨年より大幅に減少(75->15)した以外は、例年と大きな違いは見られなかった。
チョウチョウウオ類が-5mのように多くなかったのは、サンゴの群体形状の違いで小さなチョウチョウウオ類が隠れられるような場所が
多くないためと思われる。

参加者の感想
南部 西崎サンゴのリーフチェックに参加して Matsushita
西崎サンゴはサンゴの北端に位置するなかで大きな群生であることは聞いていたが、こんなに大きな群生であるとは知らなかった。串本に負けない群生だった。初めての参加で魚類調査班のクロホシイシモチ、ソラスズメダイやイサキなど担当になり、これらは非常に個体数が多く大変やり甲斐があった。目を白黒させながら数えました! これでイサキが回ってきたら嬉しい悲鳴になっていたのではと思います。
数百匹を数えるのは大変。群れは移動するし、サンゴの下に隠れているしで数えることに集中して、他の生物やサンゴをじっくり観察する暇が無いほど忙しく、少し残念でした?
終了後、科学者さんによる調査結果の総評を聞いて、継続的なデータは必要とするが環境は急速にどんどん悪い方向に向かってるんだろうなと思ってしまった。損傷を受けているサンゴが全体の1/3にも登っていることや、フクロノリが増えていることなどを教えてくださった。また着実に温帯域が魚類の分布が亜熱帯科しつつあることなどを聞いて、これが人間の愚かな行動によるところの原因でなっているんだろうなあ・・・と、アルコールが回りつつある頭で残念な気持ちになってしまいました。
全体を通じて感じたことは、リーフチェックは気さく楽しいダイビングの一つだなと思った。堅苦しく考える必要は全くなく、FUNダイブとして機会があれば皆さんも体験して見てください。今までとは違ったダイビングの楽しさ、新しい発見や人間と自然の関わり方を考える良い機会になると思います。また次回も時間と機会があれば参加したいと思います。
参加者の感想 スタッフAさん
始めは調査と言う事で、難しいそうなイメージがあったけど、やってみれば気軽に簡単にできました。遊びのダイビングだけではなく、年に1回でもこのような事に参加する事も良いのではないでしょうか?他人がやるのではなく、個人個人が考えて行動する事が大切だと改めて強く思いました。参加して頂いた方からも、こういったダイビングもおもしろい・楽しかったと言う感想が多かったです。大切な資源を守っていく事が、これからのダイビングにも良い影響を与えてくれるのではないかと思います。








第1回 南部・田辺エリア(沖ノ島)リーフチェック 2004.05.29-30
ポイント『ニシザキサンゴ』で始めてRCが実施されます。とてもサンゴのキレイなポイントで、ダイバーにも人気です。このポイントは大切にしていきたいと考え、継続的にモニタリングしていきたいと企画しました。何事も始めの1歩から!ご協力よろしくお願いします。
【参加者の声;もり2】
「なにぃ!南部で初めてぇ?!」。初物好きのおいやんは「初めて」のキーワードにコロリとやられ、3月から月一でリーフチェック(RC)に向けて勉強会 に参加。回を重ねるたびに「いや~、こんなんホンマにできるんやろかぁ~しんじられへ~ん」って不安いっぱいで本番を迎えました。ですが実際やってみる と。。。
自分の分担は数種類にしぼられ集中して調査できたので案外こなせたし(調査対象についてもよく勉強できたよ)、すきまを覗き込んでくまなく「探す」こと に集中していたので、いつもよりいろんなものを(自分で)見つけられて、「おもしろい!!」、こんなんもありやなぁ~って思いました。この調査データ は、サンゴを囲む生態系がどうなってきているのかを経年を追って把握するため、またその結果によっては行政に環境改善をもとめていくための基礎データと してもつかっていくということです。
今回調査の合間にニシザキサンゴをみて再認識!「このすばらしいサンゴは我々の手で守らにゃアカン!!」って。サンゴ&スズメダイ系乱舞の風景が大好きなおいやんとしては、ダイビングを末永く楽しんでいくための重要な一つの活動としてこれからも参加していきたいです。今回参加されなかった人も次回はどんどん参加してほしいと思います。やっぱ何事も経験しないと意識はあがりませんから。。。みんなで「コーラルコンシャス」なダイバーになろう!!(あれっ???なんかこのフレーズどっかで聞いたな。。。ぱくり??)
【参加者の声;やまちゃん】
リーフチェックお疲れ様でした。今回南部初という事でしたが、人数も結構集まって大々的に活動できた事に達成感と大きな意味があったのではないでしょうか!作業自体も簡単で中性浮力をそこそこ出来れば、誰でも参加出来るイベントだと思います。来年も5月末あたりにするという事なので、珊瑚や環境問題等に関心があればみなさん是非参加してもらい、貝駆除も含めてこれから、どうすれば今の海を守れるかを考えてもらえれば幸いかなと思います。
本当にありがとうございました(@^∀^@)

★今回の調査は大変有意義なものになりました。 RC参加の皆さん 本当にお疲れ様でした。
29日はFUNダイブ+以前に南部鹿島ビーチに移植したサンゴの調査ダイブ、30日のRCの2日間合計で312分潜りました。充分な潜水時間(笑う)でした。来年も宜しくお願い致します。
●もし、秋の時期にRCを実施できれば、タカサゴやら南方系のチョウチョウウオがたくさん見ることができますし季節による生物量の確認もできると思います。今年は早くもノコギリダイ5固体、浅瀬には小ぶりのタカサゴも確認できました。
●ショックなことは30cmクラスのオニヒトデを発見したことです。すぐに水中で駆除しましたが、充分に産卵できるサイズですし今後が心配です。 串本にも今年浅瀬に現れたそうです。 すさみには以前から1ダイブに1回は確認できるほど生息しています。なぜ紀伊半島の中間地点の「すさみ」に生息しているかは疑問となるところですが。 白浜沖では今年に入って10cm程度のオニヒトデが、沢山漁師の網にかかるそうです。どれくらい沢山かはわかりません。
●ニシザキサンゴは沖縄に例えるならば、日本北端の石西礁湖でしょうか。今回潜ったところの近辺にも沢山のサンゴが群生しております。ダイバーが殆ど入っていない所もいっぱいあると漁師さんにも聞いていますし調査すれば、まだまだ多くの群生地を発見できると思います。
●この地域を大切に守って、保全していくことが、我々ダイバーにしかできない活動と思っております。 今後も皆さんの協力のもと、しっかりとデータを蓄積するとともに、このすばらしい北端のサンゴ群落を保全していきます。
●サンゴ食巻貝駆除やオニヒトデ駆除、そして、例えば、「鹿島ビーチにサンゴを再生」を、まさに人海戦術で短期間(1日~2日)に再生することができればと考えております。 サンゴ移植については様々な見解があり意見が分かれるところではありますが、実現は充分可能です。 白浜にも移植しましたが条件的にも悪かったにも関わらずしっかりと付いています。
●早急に実施すべきことは、サンゴ生息域を特定することと思います。 生息域の水中地図を完成させ、範囲を明確にする。
●次に、サンゴ食巻貝駆除やオニヒトデ駆除を実施しつつ定点観察、さらにRCも加えてサンゴ群落の環境をモニタリングしていくことができればと考えています。皆様のご協力宜しくお願い致します。nakaya






2004年3月6,7日:石垣島リーフチェック参加しました!
調査というとなんだか難しそうだけど、一般ダイバーが気軽に参加できるような方法になっていました!
難しく考えず、まずは参加することが大切!と感じました。年に何回かは、こういったダイビングをすることも良いのではないでしょうか?いつも楽しませてくれる海に感謝の気持ちを込めていかがでしょうか。普段と違った潜り方をして楽しかったという感想が多かったです。その後のファンダイブにも良い影響を与えてくれそうです。
【サンゴ群集の生物多様性調査(リーフチェック等)】
サンゴ群集の生物多様性調査は主にサンゴ群集に対する影響を観察して、サンゴ礁保護に結びつけていく為のモニタリング調査です。
・サンゴ礁を身近な視点で捉え、サンゴ礁の価値について考え、その保護に寄与していく
・環境保護の必要性や活動を社会的にアピールしていく
この2点が活動の主な目的です。
そのために、多くのレジャーダイバーにとって、・・・(続きを読む)
わかりやすく、楽しみながら実施できることが、リーフチェックの最も大切なコンセプトです。
リーフチェックは無条件にサンゴ礁を保護しよう、という活動ではありません。サンゴ礁を継続的に利用していく道を見つけ、続けていくためにリーフチェックがあります。
・サンゴ礁の健康状態を診るために、
・調査地にすむ生物(魚類・無脊椎動物)の個体数を調べ、
・サンゴの表面がどうなっているか(底質調査)、
・人為的な破壊やゴミ、サンゴの病気白化などを調査します。
・和歌山県の田辺白浜海域でのリーフチェックは、世界共通の標準調査対象種に加え、海域の特徴的代表種も追加し調査しています。
・海底に100m範囲のメジャーを引っ張りその周りの調査対象種や底質の状況を調べます。
・これらのデータは全世界から集められ、アメリカ本部で集約、統計データとして、様々なサンゴ礁保護に役立てられます。
世界的に統一した方法でサンゴ礁とそこにすむ生物を調査するのがリーフチェック!科学的に客観的に自然を捉えていくモニタリングは、将来を見通した積極的な自然保全に有効な手段です。ダイバーにしかできないボランティアです。難しく考えず、まずは参加してみよう!
サンゴ群集の生物多様性調査員(リーフチェック等)募集

・サンゴ群集の生物多様性調査(リーフチェック等)は主にサンゴ礁にたいする人為的な影響を観察してサンゴ礁保護に結び付けていく為のモニタリング調査です。
・サンゴ礁の生態系の状態を見るために、調査地にすむ生物(魚類・無脊椎動物)の個体数、サンゴの表面がどうなっているか(底質調査)、人為的な破壊やゴミ、サンゴの病気白化などを調査します。 魚類と無脊椎生物は世界共通の標準調査対象種が決められています。
・海底に100m範囲のメジャーを引っ張りその周りの調査対象種の数を調べます。
・調査は一般のレジャーダイバーでも容易にできるようプログラムされています。難しく考えず気軽に参加頂けますと幸いです。
令和7年度の調査日程
令和7年度の公示調査は終了しました。
※調査には参加条件(例えば潜水士免許・調査経験など)あります。詳しくは「お問い合わせフォーム」よりお問合せ下さい。
基本スケジュール(海象等により出船時間等が変更になる場合があります)
基本スケジュールA
8:00集合 現地ダイビングサービス(田辺市内・白浜町内)
8:30 調査担当確認、器材等準備、ボート乗船
10:00 出船 調査ダイブ1
11:30 帰港
昼食等(自分でご用意下さい)
13:00 出船 調査ダイブ2
14:30 帰港、調査データシート回収、器材洗浄、シャワー等
15:00 調査ヒヤリング、データ発表等
16:30 修了・解散
※詳しくは基本スケジュールBも参照下さい。
基本スケジュールB
基本スケジュールBは、 ・・・(続きを読む)
8:00集合 現地ダイビングサービス(田辺市内・白浜町内)
8:15~調査担当確認
9:00~準備、ボート乗船
9:30 出港
9:45 ポイント到着
9:50 ライン設置
10:05 魚類調査班エントリー(ライン設置15分後)
10:20 無脊椎調査班エントリー(魚類EN15分後)
10:35 底質調査班エントリー(無脊椎EN15分後)
11:15 調査終了、ライン回収、データシート点検、GPS測定
11:30 港着
昼食
13:00 出港
13:15 ポイント到着
13:20 ライン設置
13:45 魚類調査班エントリー(ライン設置15分後)
14:00 無脊椎調査班エントリー(魚類EN15分後)
14:15 底質調査班エントリー(無脊椎EN15分後)
15:00 調査終了、ライン回収、データシート点検、GPS測定
15:30 港着~機材片付け
16:00 データ集計
16:30~データ発表、TS(チーム科学者)からのコメント、TL(チームリーダ)からの総評など
17:00 修了
準備するもの
・遊びのダイビングと同様の準備をして下さい。水着、着替え、器材等です。
・ダイビング器材のレンタル(有料)もございます。お申込み時にご要望下さい。
・調査で使用する器材は、弊社で準備致します。
参加資格と費用
★令和7年度は、生物多様性調査は調査経験者のみとし、未経験者は参加できません。ご了承下さい。
1.調査未経験者は、
・・・(続きを読む)
(1)レクチャ受講(プレ調査ダイブと同日に実施)
・レクチャは30分程度実施します。
・魚類や海洋生物の図鑑等持っている方は持参下さい。
(2)プレ調査ダイブ(実習1ビーチダイブ)への参加
・プレ調査ダイブは、サンゴ群集に実際に潜り、調査の実習を実施します。
・同日に(1)項のレクチャも実施します。
☆プレ調査ダイブ参加費用:15,000円(税込、器材・個人的経費等含まない)
・午前9:30集合~12:00 ・午後13:00集合~15:30
※3名様参加より調整します。
2.ダイビング経験(下記事項の全てに該当する方)
・潜水士資格保有者 ・アドバンスCカード以上の保持者 ・50ボートダイブ以上の経験 ・健康な方 ・オウンリスクで参加できる方 ※自分で潜れない方は参加できません。
3.調査一般参加費用:通常の遊びのダイビング費用程度
・費用に含むもの(乗船料2回、タンク2本、調査道具1式)
・費用に含まれないもの(上記以の個人的な経費で飲食代、交通費、宿泊費、器材等)
※参加申し込みは「お問い合わせフォーム」から送信下さい。